Apicomplexa
Apicomplexa ili Apicomplexia (lat. apex = gore + complexus = kompleks – odnosi se na skup organela u sporozoitu) je veliko koljeno parazitskih Alveolata. Većina njih ima jedinstven oblik organela sa plastidima zvanim apikoplasti i apikalnu složenu strukturu. Organela je adaptacija koju apikoplekse koriste pri penetraciji u ćeliju domaćina.
| Apicomplexa (Koljeno parazitskih Alveolata) | |
|---|---|
 Zbirka apopleksnih životinja | |
| Sistematika | |
| Koljeno | Apicomplexa |
| |
Apicomplexa su jednoćelijska i sporne. Sve su vrste obligatne endoparazitske životinje, osim u rodu Nephromyces, simbionti morskih životinja, prvobitno klasificirane kao gljive Chytridae. Pokretne strukture, kao što su bičevi ili pseudopodje prisutne su samo u određenim fazama gameta[1][2] Apicomplexa su raznolika grupa koja uključuje organizme poput kokcidija, gregarina, piroplazme, hemogregarina i plazmoji.
Apicomplexa izazivaju bolesti, kao što su:
- Babesioza (Babesia)
- Malarija (Plasmodium)
- Kriptosporidioza (Cryptosporidium parvum)
- Ciklosporijaza (Cyclospora cayetanensis)
- Cistoizosporijaza (Cystoisospora belli (bivša "Isospora Belli"))
- Toksoplasmoza (Toxoplasma gondii)[3].
Apicomplexa obuhvata većinu grupe koja se nekada zvala Sporozoa, grupu parazitskih "protozoa" ", uglavnom bez bičeva, treplji ili pseudopodija. Međutim, većina je pokretna upotrebom klizajućih mehanizama, koji počiva na statičkoj adheziji malih motornih miozina. Ostale glavne linije bile su Ascetosporea (sada u Rhizaria), Myxozoa (sada je poznato da su visoko izvedene knidarijske životinje), i Microsporidia (za koji se sada zna da potiču od gljiva). Naziv Sporozoa uzima se ponekad kao sinonim za Apicomplexa, ili povremeno kao podskup.[4]
Opis
uredi
2 – konoid,
3 – mikronema,
4 – sekrecijaka organela,
5 – jedro,
6 – jedarce,
7 – mitohondrija,
8 – stražnji prsten,
9 – alveole,
10 – Golgijev aparat,
11 – mikropore

Koljeno Apicomplexa sadrži sve eukariote sa grupom struktura i organela koje se zajedno nazivaju apikalni kompleks. Ovaj kompleks sastoji se od strukturnih komponenti i sekrecijske organele koje su potrebne za invaziju na domaćinske ćelije tokom parazitskih faza životnog ciklusa. Apicomplexa imaju složene životne cikluse koji uključuju nekoliko faza i obično prolaze i aseksualne jedinke i spolnu replikaciju. Sve Apicomplexa su, u nekom dijelu životnog ciklusa obligatni paraziti, pri čemu neki parazitiraju na dva odvojena domaćina u aseksualnim i seksualnim fazama.
Pored sačuvanog apikalnog kompleksa, Apicomplexa su i morfološki raznoliki. Različiti organizmi unutar Apicomplexa, kao i različiti životni stadiji određenih apikompleksa, mogu značajno varirati u veličini, obliku i subćelijskoj strukturi. Kao i drugi eukarioti, Apicomplexa imaju jedro , endoplazmatski retikulum i Golgijev kompleks. Obično imaju jednu mitohondriju, kao i drugi izvedeni endosimbionati organel zvanu apikoplast koja ima zasebni 35-kilobazni kružni genom (s izuzetkom vrsta roda Cryptosporidium i Gregarina niphandrodes, kojima nedostaje apikoplast). Svi članovi ovog tipa imaju infektivni stadij – sporozoit – koji u apikalnom kompleksu ima tri različite strukture. Apikalni kompleks sastoji se od skupa spiralno raspoređenih mikrotubula s (konoidom), sekretorna tijela i jedan ili više polarni prstenova. Mogu imati i dodatna vitka sekretorna tijela koja su gusta od elektrona (mikroneme), okružena sa jednim ili dva polarna prstena. Ova struktura daje naziv taksonu. Daljnja skupina sfernih organela raspoređena je u cijeloj ćeliji, a nije lokalizirana u apikalnom kompleksu i poznate su kao guste granule. Obično imaju srednji promjer oko 0,7 µm. Izlučivanje sadržaja gustih granul događa se nakon invazije parazita i lokalizacije unutar parazitoforne vakuole i traje nekoliko minuta.
- Bičevi nalaze se samo u pokretnim gametima. Usmjereni prema naprijed i razlikuju se u broju (obično jedan do tri).
- Bazna tijela su takošer prisutna. Iako hemosporidijei piroplazmidi, u baznim tijelima imaju normalne trostruke mikrotubule, kokcidije i gregarine imaju devet jednostrukih.
- Mitohondrije imaju cjevaste kriste.
- Centriole, hloroplasti, izbacive organele i inkluzije nemaju.
- Ćelija je okružena pelikulom, sa tri membranska sloja (alveolarna struktura) koja prodire kroz mikropore.
Replikacija:
- Mitoza je obično zatvorena, s unutarjedarnim diobenim vretenom; kod nekih vrsta otvorena je na polovima.
- Ćelijska dioba obično je shizogonija.
- Mejoza javlja se u zigotima.
Mobilnost: Apikompleksa imaju jedinstvenu sposobnost klizanja koja im omogućava prolazak kroz tkiva i ulaz u i napuštanje ćelija domaćina. Ova sposobnost omogućena je primjenom adhezija i malih statičkih motornih miozina. Ostale karakteristike koje su zajedničke ovom tipu su nedostatak cilija, seksualna reprodukcija, upotreba mikropora za uzimanje hrane i proizvodnja oocista koje sadrže sporozoite kao zarazni oblik. Čini se da su u ovom tipu transpozoni rijetki, ali su identificirani u rodovima Ascogregarina i Eimeria.
Životni ciklus
uredi
Većina predstavnika apikompleks ima složen životni ciklus, koji uključuje i aseksualnu i seksualnu reprodukciju. Obično se domaćin inficira aktivnom invazijom parazita (slično entozi), koji se dijele i proizvode sporozoite koji ulaze u njegove ćelije. Na kraju se ćelije rasprsnu, oslobađajući merozoite, koji inficiraju nove ćelije. To se može dogoditi nekoliko puta, sve dok se ne naprave gamonti, koji formiraju gamete koje se spajaju kako bi stvorili nove ciste. Međutim, u ovom osnovnom obrascu pojavljuju se mnoge varijacije, a mnogi Apicomplexa imaju više od jednog domaćina.
Apikalni kompleks uključuje vezikule nazvane roptrije i mikroneme, koji se otvaraju u prednjem dijelu ćelije. Sekretini enzimi omogućavaju parazitu da uđe u druge ćelije. Vrh je okružen trakom mikrotubula, nazvanom polarnim prstenom, a među konoidima je i lijevak tubulinskih proteina koji se nazivaju konoidi. Preko ostatka ćelije, osim umanjenih usta zvanih mikropore, membrana se oslanja na vezikule zvane alveole, tvoreći polukrutu pelikulu.
Prisustvo alveola i drugih osobina svrstava Apicomplexa u grupu koja se zove alveolata. Nekoliko srodnih alveola, poput Perkinsus i Colpodella, imaju strukture slične polarnom prstenu i ranije su bile ovdje uključene, ali čini se da su većina bliži srodnici dinoflagelata . Vjerovatno su slične zajedničkom pretku dviju skupina.
Još jedna sličnost je da mnoge ćelije apikopleksa sadrže jedan plastid, zvani apikoplast, okružen s tri ili četiri membrane. Smatra se da njegove funkcije uključuju zadatke poput biosinteze lipida i hema, a čini se da im je to potrebno za opstanak. Općenito, smatra se da plastidi imaju zajedničko porijeklo s hloroplastima dinoflagelata, a dokazi upućuju na porijeklo iz crvenih, a ne iz zelenih algi.[5][6]
Podgrupe
urediApikompleksa imaju vrste tri grupe – kokcidijska, gregarinska i hemosporidna. Izgleda da su kokcidi i gregarini srodnički povezani. Perkinsus, iako se nekad smatrao članom Apicomplexa, premješten je u novi takson – Perkinsozoa.[7]
Gregarine
uredi

Gregarine su obično paraziti anelida, artropoda i školjki. Često se nalaze u crijevima svojih domaćina, ali mogu upasti i u ostala tkiva. U tipskom životnom ciklusu gregarina, unutar ćelije domaćina, trofozoit se razvija u shizont. Zatim se shizogonijom dijeli na merozoite. Merozoiti se oslobađaju razgradnjom ćelije domaćina i upada u ostale ćelije. U nekom dijelu životnog ciklusa apikompleksa formiraju se gametociti. Oni se oslobađaju razgradnjom ćelija domaćina, koje se grupiraju. Svaki gametocit formira više gameta. Gume se stapaju s drugom i tvore oociste. Oociste preuzimaju domaćini da ga preuzmu novi domaćini.
Kokcidije
urediKokcidije su općenito paraziti kičmenjaka s. Kao i gregarine, oni su obično paraziti epitelnih ćelija crijeva, ali mogu zaraziti i druga tkiva.
Životni ciklus kokcija uključuje meronije, gametogonije i sporogonije. Iako je sličan onom gregarinama, razlikuje se u formiranju zigota. Neki trofozoiti se povećavaju i postaju makrogameti, dok se drugi opetovano dijele i tvore mikrogamete (anizogamija). Mikrogameti su pokretni i mgu doći do makrogamatima da ih oplode. Oplođeni makrogamet formira zigot koji se pretvara u oocistu koja se normalno oslobađa iz tijela. Shizigija, kada se pojavi, uključuje izrazito neskladne gamete. Životni ciklus je obično sa haploidima, s jedinom diploidnom fazom u zigotu, a koja je obično kratkotrajna.
Glavna razlika između kokcidija i gregarina je u gamantima. Kod kokcidija su to male, unutarćelijske i bez epimera ili mikrokorona. U gregarinima su to velike, vanćelijske, sa epimeritima ili mikronima. Druga razlika između kokcidija i gregarina također je u gamontima. U kokcidijama pojedinačni gamont postaje makroametocit, dok u gregarina gamonti stvaraju višestruke gametonite.
Haemosporidia
uredi
Hemosporidije imaju složenije životne cikluse, koji se razlikuju između člankonožaca i domaćina kičmenjaka. Trofozoit parazitira u eritrocitima ili drugim tkivima kičmenjaka domaćina. Mikrogameti i makrogameti se uvijek nalaze u krvi. Gamete preuzima vektorski insekt tokom sisanja krvi. Mikrogamete migriraju u crijevima insekata i spajaju se s makrogametima. Oplođeni makrogamat tada postaje ookineta koja prodire u tijelo vektora. Ookinete se zatim pretvaraju u oociste i dijele se u početku mejotski, a zatim mitozom (haplontni životni cikluso) da bi se stvorio sporozoit. Sporozoiti bježe iz oociste i migriraju unutar tijela vektora do pljuvačnih žlijezda gdje se ubrizgavaju u organizam novog kičmenjaka domaćina kada se vektor insekata ponovno hrani.
Ekologija i distribucija
urediMnogi paraziti apikompleksa važni su uzročnici bolesti ljudi i domaćih životinja. Za razliku od patogenih bakterija, apikoplekski paraziti su eukariotski i dijele mnoge metaboličke puteve sa svojim životinjskim domaćinima. Zbog toga je terapijski ciljni tretman izuzetno težak - lijek koji šteti apikopleksnom parazitu vjerovatno će štetiti i čovjeku domaćinu. Dosad nije bila dostupna efikasna vakcina za većinu bolesti izazvanih tim parazitima. Biomedicinska istraživanja ovih parazita izazovna su jer je često teško, ako ne i nemoguće, održavati žive kulture parazita u laboratoriji i genetički manipulirati tim organizmima. Posljednjih godina nekoliko vrsta apikopleksa odabrano je za sekvenciranje genoma. Dostupnost genomskih sekvenci pruža novu priliku naučnicima da saznaju više o evoluciji i biohemijskim osobenostima ovih parazita. Pretežni izvor tih genskih informacija je baza EuPathDB Plasmodium. Baza podataka koja uključuje eksperimentalne i računarske podatke za kokcidije (ToxoDB), piroplazme (PiroplasmaDB) i vrsta roda Cryptosporidium (CryptoDB). Jedan od mogućih ciljeva za pripravljanje lijekove je da djeluje na plastid, a čini se da postojeći lijekovi poput tetraciklina, koji su efikasni protiv apikopleksa, djeluju i protiv plastida.
Mnogi kokcidiomorfi imaju srednjeg domaćina, kao i primarnog domaćina, a evolucija izbora domaćina odvijala se na različite načine i u različitim vremenima u tim skupinama. Za neke kokcidiomorfe izvorni domaćin postao je posredni domaćin, dok je za druge postao konačni. U rodovima Aggregata, Atoxoplasma, Cystoisospora, Schellackia i Toxoplasma, original je sada definitivan, dok je u rodovima Akiba , Babesiosoma, Babesia , Haemogregarina, Haemoproteus, Hepatozoon, Kariolysus, Leucocytozoon , Plasmodium, Sarcocystis i Theileria, izvorni domaćini su sada posredni.
Slične strategije za povećanje vjerovatnoće prenošenja razvijale su se u više rodova. Ciste polienergidnih oocista i tkiva nalaze se u predstavnicima redova Protococcidiorida i Eimeriida. Hipnozoit se nalaze u Karyolysus lacerata i u većini vrsta plazmodijum; transovarijalni prijenos parazita događa se u životnim ciklusima rodova Karyolyzus i Babesia.
Pretpostavka je da se horizontalni transfer gena u evoluciji ovog koljena dogodio relativno rano, prenošenjem histona H4 u lizinu 20 (H4K20), kao modifikator KMT5A ( Set8), od životinje domaćina do pretka apikopleksana. Drugi gen – H3K36 metiltransferaza (Ashr3) – može se prenijeti i horizontalno.
Krvni paraziti
urediUnutar Apicomplexa nalaze se paraziti:
- podred Adeleorina - osam rodova
- podred Haemosporina - svi rodovi u podrdu
- podred Eimeriorina - dva roda ( Lankesterella i Schellackia )
Unutar grupe Adelorine nalaze se vrste koje inficiraju beskičmenjake i druge koje inficiraju kičmenjake. Eimeriorina – najveći podred u ovom tipu – životni ciklus uključuje i seksualnu i aseksualnu fazu. Aseksualne faze reproduciraju se shizogonijom. Muški gametocit stvara veliki broj gameta, a zigot stvara oocist, što je infektivni stadij. Većina je monoksena (inficira samo jednog domaćina), ali nekoliko ih je heteroksenih (životni ciklus uključuje dva ili više domaćina). Još uvijek raspravlja se o broju porodica u ovom podredu, pri čemu je broj porodica između jedne i 20, ovisno o autorima, a broj rodova između 19 i 25.
Evolucija
urediSvi članovi ovog koljena parazitiraju i razvijaju se od slobodnoživećeg pretka . Pretpostavlja se da je ovaj način života evoluirao u vrijeme divergencije dinoflagelata i apikompleksa..[8][9] Procjenjuje se da je njihova dalja evolucija trajala oko 800 miliona godina.[10] Smatra se da su, od postojećih kladusa, najstariji arhigregarini.[11] Ti filogenetski odnosi rijetko su proučavani na nivou podrazreda. Haemosporidije su povezane sa gregarinama, a piroplazme i kokcidije su sestrinske grupe. Čini se da su Haemosporidia i piroplazma sestrinski kladusi i više su vezani za kokcidije nego za gregarine.[12]
| Apicomplexa |
| ||||||||||||||||||||||||||||||
Taksonomija
urediHistorija
urediPrve Apicomplexa vidio je Antonie van Leeuwenhoek, koji je 1674. godine u žučnom mjehuru zeca ugledao vjerovatno oociste "Eimeria stiedae". Dufour je 1828. godine imenovao prvu vrstu koljena, "Gregarina ovata" "u crijevima uholadže. On je mislio da su oni posebna grupa povezana sa trematodama, u to vrijeme uključenim u Vermes. Od tada ih je mnogo više identificirano i imenovano. Tokom 1826-1850. imenovane su 41 vrste i šest rodova Apicomplexa. U periodu 1951–1975 dodane su 1873 nove vrste i 83 nova roda.
Stariji takson Sporozoa, uključen u Protozoa, stvorio je Leuckart 1879. Kroz historiju su sa sadašnjim Apicomplexa grupirale mnoge nepovezane grupe. Naprimjer, Kudo (1954) u sporozoa uključuje vrste grupe Ascetosporea (Rhizaria), Microsporidia (Fungi), Myxozoa (Animalia) i Helicosporidium (Chlorophyta), dok je Zierdt (1978.) uključio rod Blastocystis (Stramenopiles). Dermocystidium se također smatran sporozoom. Nisu sve ove grupe imale spore, ali sve su bile parazitske. Međutim, i drugi parazitski ili simbiotski jednoćelijski organizmi,ako su imali bičeve, također su bili uključeni u protozojske skupine izvan Sporozoa (Flagellata, Ciliophora i Sarcodina) (npr., mnoge Kinetoplastida, Retortamonadida, Diplomonadida, Trichomonadida, Hypermastigida), cilija (npr. Balantidium) ili pseudopodije (npr. Entamoeba, Acanthamoeba,Naegleria). Ako imaju ćelijske zidove, takođe bi mogli biti uključene u biljno carstvo između bakterija ili kvasca.
Sporozoa se više ne smatra biološki valjanim taksonom i upotreba tog termina je obeshrabrena, iako ga neki autori još uvijek koriste kao sinonim za Apicomplexa. U novije vreme su druge grupe bile isključene iz Apicomplexa, npr. Perkinsus i Colpodella (sada u Protalveolati). Polje razvrstavanja Apicomplexa je u toku i klasifikacija se mijenjala tokom svih godina otkako je 1970. formalno imenovani takson.
Podjela
urediDo 1987. godine dovršeno je sveobuhvatno istraživanje ovog taksona: ukupno je imenovano 4516 vrsta i 339 rodova. Sastojali su se od:
- Razred Conoidasida
- Podrazred Gregarinasina pp
- Red Eugregarinorida, sa 1624 imenovane vrste i 231 rodom
- Podrazred Coccidiasina p.p
- Red Eucoccidiorida p.p
- Podred Adeleorina p.p
- Grupa Hemogregarin s 399 vrsta i četiri roda
- Podred Eimeriorina, sa 1771 vrstom i 43 roda
- Podred Adeleorina p.p
- Red Eucoccidiorida p.p
- Podrazred Gregarinasina pp
- Razred Aconoidasida
- Red Haemospororida, sa 444 vrste i devet rodova
- Red Piroplasmorida, sa 173 vrste i 20 rodova
- Ostale manje grupe izostavljene gore, sa 105 vrsta i 32 roda
Iako je izvršena znatna revizija ovog taksona (pored Haemosporidia sada ima 17 rodova, a ne 9), ovi brojevi su verovatno još uvek približno tačni.
- Vrste
- Uzročnik piroplasmoza
- Uzročnici babezioza
- Cryptosporidies, uzročnici kriptosporidioza
- Coccidies, uzročnici kokcidioza
- Eimeria ahsata
- Eimeria acervulina
- Eimeria bovis
- Eimeria brunetti
- Eimeria columbarum
- Eimeria crandallis
- Eimeria falciforme
- Eimeria flavescens
- Eimeria labbeana
- Eimeria maxima
- Eimeria meleagridis
- Eimeria mitis
- Eimeria necatrix
- Eimeria ninakohlyakimovae
- Eimeria perforans
- Eimeria praecox
- Eimeria sciurorum
- Eimeria stiedae
- Eimeria tenella
- Eimeria zurnii
- Gregarina
- Monospora
- Uzročnici neosporoza
- Plasmodium
Razni plazmodiji
- Plasmodium achiotense
- Plasmodium achromaticum
- Plasmodium acuminatum
- Plasmodium adunyinkai
- Plasmodium aegyptensis
- Plasmodium aeuminatum
- Plasmodium agamae
- Plasmodium anasum
- Plasmodium anomaluri
- Plasmodium arachniformis
- Plasmodium ashfordi
- Plasmodium atheruri
- Plasmodium aurulentum
- Plasmodium australis
- Plasmodium attenuatum
- Plasmodium azurophilum
- Plasmodium balli
- Plasmodium bambusicolai
- Plasmodium basilisci
- Plasmodium beebei
- Plasmodium beltrani
- Plasmodium berghei
- Plasmodium bertii
- Plasmodium bigueti
- Plasmodium biziurae
- Plasmodium booliati
- Plasmodium bouillize
- Plasmodium bowiei
- Plasmodium brodeni
- Plasmodium brasilianum
- Plasmodium brasiliense
- Plasmodium brumpti
- Plasmodium brucei
- Plasmodium brygooi
- Plasmodium bubalis
- Plasmodium bucki
- Plasmodium capistrani
- Plasmodium cathemerium
- Plasmodium cephalophi
- Plasmodium cercopitheci
- Plasmodium chabaudi
- Plasmodium chalcidi
- Plasmodium chiricahuae
- Plasmodium circularis
- Plasmodium circumflexum
- Plasmodium clelandi
- Plasmodium cordyli
- Plasmodium cnemaspi
- Plasmodium cnemidophori
- Plasmodium coatneyi
- Plasmodium coggeshalli
- Plasmodium colombiense
- Plasmodium columbae
- Plasmodium corradettii
- Plasmodium coturnixi
- Plasmodium coulangesi
- Plasmodium cuculus
- Plasmodium cyclopsi
- Plasmodium cynomolgi
- Plasmodium diminutivum
- Plasmodium diploglossi
- Plasmodium dissanaikei
- Plasmodium divergens
- Plasmodium dominicana
- Plasmodium draconis
- Plasmodium durae
- Plasmodium egerniae
- Plasmodium elongatum
- Plasmodium eylesi
- Plasmodium fabesia
- Plasmodium fairchildi
- Plasmodium falciparum
- Plasmodium falconi
- Plasmodium fallax
- Plasmodium fieldi
- Plasmodium fischeri
- Plasmodium foleyi
- Plasmodium formosanum
- Plasmodium forresteri
- Plasmodium floridense
- Plasmodium fragile
- Plasmodium galbadoni
- Plasmodium garnhami
- Plasmodium gallinaceum
- Plasmodium giganteum
- Plasmodium giovannolai
- Plasmodium girardi
- Plasmodium gonderi
- Plasmodium georgesi
- Plasmodium gologoense
- Plasmodium gracilis
- Plasmodium griffithsi
- Plasmodium guangdong
- Plasmodium gundersi
- Plasmodium guyannense
- Plasmodium heischi
- Plasmodium hegneri
- Plasmodium hermani
- Plasmodium heteronucleare
- Plasmodium hexamerium
- Plasmodium holaspi
- Plasmodium huffi
- Plasmodium hylobati
- Plasmodium icipeensis
- Plasmodium iguanae
- Plasmodium inconstans
- Plasmodium inopinatum
- Plasmodium inui
- Plasmodium japonicum
- Plasmodium jefferi
- Plasmodium jiangi
- Plasmodium josephinae
- Plasmodium joyeuxi
- Plasmodium juxtanucleare
- Plasmodium kempi
- Plasmodium knowlesi
- Plasmodium kentropyxi
- Plasmodium lacertiliae
- Plasmodium lainsoni
- Plasmodium landauae
- Plasmodium leanucteus
- Plasmodium lemuris
- Plasmodium lepidoptiformis
- Plasmodium lionatum
- Plasmodium lophurae
- Plasmodium loveridgei
- Plasmodium lutzi
- Plasmodium lygosomae
- Plasmodium mabuiae
- Plasmodium mackerrasae
- Plasmodium maculilabre
- Plasmodium maior
- Plasmodium majus
- Plasmodium malariae
- Plasmodium marginatum
- Plasmodium matutinum
- Plasmodium melanipherum
- Plasmodium mexicanum
- Plasmodium michikoa
- Plasmodium minasense
- Plasmodium morulum
- Plasmodium murinus
- Plasmodium necatrix
- Plasmodium nucleophilium
- Plasmodium octamerium
- Plasmodium odocoilei
- Plasmodium osmaniae
- Plasmodium ovale
- Plasmodium paddae
- Plasmodium papernai
- Plasmodium paranucleophilum
- Plasmodium parvulum
- Plasmodium pedioecetii
- Plasmodium pelaezi
- Plasmodium percygarnhami
- Plasmodium pessoai
- Plasmodium petersi
- Plasmodium pifanoi
- Plasmodium pinotti
- Plasmodium pinorrii
- Plasmodium pitheci
- Plasmodium pitmani
- Plasmodium polare
- Plasmodium pulmophilum
- Plasmodium reichenowi
- Plasmodium relictum
- Plasmodium rhadinurum
- Plasmodium rhodaini
- Plasmodium robinsoni
- Plasmodium rouxi
- Plasmodium sandoshami
- Plasmodium sasai
- Plasmodium saurocaudatum
- Plasmodium schweitzi
- Plasmodium scorzai
- Plasmodium semiovale
- Plasmodium semnopitheci
- Plasmodium shortii
- Plasmodium silvaticum
- Plasmodium simium
- Plasmodium tanzaniae
- Plasmodium tenue
- Plasmodium tejerai
- Plasmodium telfordi
- Plasmodium tomodoni
- Plasmodium torrealbai
- Plasmodium traguli
- Plasmodium tribolonoti
- Plasmodium tropiduri
- Plasmodium tumbayaensis
- Plasmodium tyrio
- Plasmodium uilenbergi
- Plasmodium uluguruense
- Plasmodium uncinatum
- Plasmodium uzungwiense
- Plasmodium watteni
- Plasmodium wenyoni
- Plasmodium vacuolatum
- Plasmodium vastator
- Plasmodium vaughani
- Plasmodium vautieri
- Plasmodium vinckei
- Plasmodium vivax
- Plasmodium volans
- Plasmodium voltaicum
- Plasmodium yoelii
- Plasmodium youngi
- Plasmodium zonuriae
- Theileria
- Theileria annulata (uzročnik teilerioza)
- Theileria parva (uzročnik "groznice istočnih obala")

- Toksoplazma, uzročnik toksoplasmoza
- Fosili
Galerija
uredi-

-

-
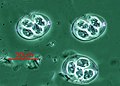 Cista Eimeria maxima (Coccidiasina)
Cista Eimeria maxima (Coccidiasina) -
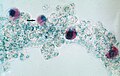
 Cista Sarcocystis hominis (Coccidiasina)
Cista Sarcocystis hominis (Coccidiasina) -

-

-

-

-

-









Također pogledajte
urediReference
uredi- ^ Jadwiga Grabda (1991). Marine fish parasitology: an outline. VCH. str. 8. ISBN 978-0-89573-823-3.
- ^ Saffo M. B.; McCoy A. M.; Rieken C.; Slamovits C. H. (2010). "Nephromyces, a beneficial apicomplexan symbiont in marine animals". Proceedings of the National Academy of Sciences. 107 (37): 16190–5. doi:10.1073/pnas.1002335107. PMC 2941302. PMID 20736348.
- ^ Stefan H.I. Kappe; et al. (januar 2004). "Apicomplexan gliding motility and host cell invasion: overhauling the motor model". Trends in Parasitology. 20 (1): 13–6. doi:10.1016/j.pt.2003.10.011. PMID 14700584.
- ^ Sibley, LDI (Oct 2010). "How apicomplexan parasites move in and out of cells". Curr Opin Biotechnol. 21 (5): 592–8. doi:10.1016/j.copbio.2010.05.009. PMC 2947570. PMID 20580218.
- ^ Patrick J. Keeling (2004). "Diversity and evolutionary history of plastids and their hosts". American Journal of Botany. 91 (10): 1481–1493. doi:10.3732/ajb.91.10.1481. PMID 21652304. Arhivirano s originala, 27. 2. 2008. Pristupljeno 19. 2. 2020.
- ^ Ram, Ev; Naik, R; Ganguli, M; Habib, S (juli 2008). "DNA organization by the apicoplast-targeted bacterial histone-like protein of Plasmodium falciparum". Nucleic Acids Research. 36 (15): 5061–73. doi:10.1093/nar/gkn483. PMC 2528193. PMID 18663012.
- ^ Norén, Fredrik; Moestrup, Øjvind; Rehnstam-Holm, Ann-Sofi (oktobar 1999). "Parvilucifera infectans norén et moestrup gen. et sp. nov. (perkinsozoa phylum nov.): a parasitic flagellate capable of killing toxic microalgae". European Journal of Protistology. 35 (3): 233–254. doi:10.1016/S0932-4739(99)80001-7.
- ^ Kuvardina ON, Leander BS, Aleshin VV, Myl'nikov AP, Keeling PJ, Simdyanov TG (novembar 2002). "The phylogeny of colpodellids (Alveolata) using small subunit rRNA gene sequences suggests they are the free-living sister group to apicomplexans". J. Eukaryot. Microbiol. 49 (6): 498–504. doi:10.1111/j.1550-7408.2002.tb00235.x. PMID 12503687.
- ^ Leander BS, Keeling PJ (august 2003). "Morphostasis in alveolate evolution". Trends Ecol Evol. 18 (8): 395–402. doi:10.1016/S0169-5347(03)00152-6.
- ^ Escalante AA, Ayala FJ (juni 1995). "Evolutionary origin of Plasmodium and other Apicomplexa based on rRNA genes". Proc. Natl. Acad. Sci. U.S.A. 92 (13): 5793–7. doi:10.1073/pnas.92.13.5793. PMC 41587. PMID 7597031.
- ^ Morrison DA (august 2009). "Evolution of the Apicomplexa: where are we now?". Trends Parasitol. 25 (8): 375–82. doi:10.1016/j.pt.2009.05.010. PMID 19635681.
- ^ Templeton TJ, Enomoto S, Chen WJ, et al. (februar 2010). "A genome-sequence survey for Ascogregarina taiwanensis supports evolutionary affiliation but metabolic diversity between a Gregarine and Cryptosporidium". Mol. Biol. Evol. 27 (2): 235–48. doi:10.1093/molbev/msp226. PMC 2877549. PMID 19778951.