Microsporidia
Microsporidia je grupa sporulirajućih jednoćelijskih parazita. Smatrane su protozoama ili protistima, ali se sada zna da su gljive[5] ili sestrinska grupa gljiva.[6] Slobodno procjenjujući, ima ih 1500 vrsta, a vjerovatno više od milion[7] do sada imenovanih.
| Microsporidia | |
|---|---|
 | |
| Sistematika | |
| Carstvo | Fungi |
| Divizija | Rozellomycota |
| Razred | Microsporidia |
| Sinonimi | |
Microsporidia su ograničene na životinje domaćine, a sve glavne grupe životinja su om domaćini. Većina zarazi insekte, ali su odgovorni i za uobičajene bolesti rakova i riba. Navedene vrste microsporidia obično zaraze jednu vrstu domaćina ili grupu blisko srodnih taksona. Nekoliko vrsta, od kojih je većina oportunističkih, inficira i ljude.
Približno 10 % vrsta čine paraziti kičmenjaka – uključujući i ljude, kod kojih mogu izazvati mikrosporidijozu. Nakon infekcije, oni na različite načine utiču na svoje domaćine i napadaju sve organe i tkiva, mada općenito specifično različite vrste mikrosporidija. Neke su vrste smrtonosne, a neke se koriste u biološkoj kontroli insekata štetočina. Parazitskaa kastracija, gigantizam ili promjena spola domaćina, potencijalni su efekti mikrosporidijskog parazitizma (kod insekata). U najsavremenijim slučajevima parazitizma mikrosporidije u potpunosti ovladaju ćelijom domaćina i kontrolirau njen metabolizam i razmnožavanje, formirajući ksenome.[8] Replikacija se odvija unutar ćelija domaćina, koji su zaraženi jednoćelijskim sporama, koje su prečnika u rasponu od 1–40 µm, što ih čini jednim od najmanjih eukariota. Microsporidije koje inficiraju sisare imaju promjer oko 1,0–4,0 μm.[9] They also have the smallest eukaryotic genomes. Ermin "microsporidium" također se koristi kao zamjena za članove incertae sedis.[10]

Morfologija
urediMikrosporidije nemaju mitohondrija, umjesto kojih imaju mitosome. Također im nedostaju strukture za kretanje, poput bičeva. Mikrosporidije stvaraju visoko otporne spore, sposobne da prežive izvan svog domaćina i do nekoliko godina. Morfologija spore korisna je u razlikovanju različitih vrsta. Spore većine vrsta su ovalne ili kruškolike, ali i šipkaste ili sferne spore nisu neuobičajene. Nekoliko rodova ima spore jedinstvenog oblika za taj rod.
Spora je zaštićena zidom koji se sastoji od tri sloja:
- vanjski elektronski gust egzospor;
- srednji, široki i naizgled nestrukturirani endospor, koji sadrži hitin;
- tanka unutrašnja plazmamembrana.
U većini slučajeva postoje dva usko povezana jedra, tvoreći "diplokarion", ali ponekad postoji samo jedan.
Prednja polovina spore sadrži aparat sličan harpunu s dugačkim navojem „polarnog vlakna“ koji je namotan u stražnjoj polovini spora. Prednji dio polarne niti je okružen "polaroplastom", lamelnom membranom. Iza polarnog vlakna nalazi se stražnja vakuola.[8]
Infekcija
urediU crijevima domaćina spora klija, stvarajući osmotski pritisak sve dok mu kruti dio zida ne ošteti na najtanjem mjestu na vrhu. Stražnja vakuola nabubri, prisiljavajući polarni filament da brzo izbaci infektivni sadržaj u citoplazmu potencijalnog domaćina. Istovremeno se materijal vlakna preuređuje tako da formira cijev koja djeluje kao hipodermna igla i prodire u epitel crijeva.
Unutar ćelije domaćina, raste sporoplazma, dijeli ili formira multinukleat plazmodija, prije stvaranja novih spora. Životni ciklus se među njima znatno razlikuje. Neki imaju jednostavan aseksualni životni ciklus,[12] dok drugi imaju složen, koji uključuje više domaćina i aseksualnih i seksualnih reprodukcija. Različite vrste spora mogu se stvarati u različitim fazama, vjerojatno s različitim funkcijama, uključujući autoinfekciju (prijenos unutar jednog domaćina).
Medicinske implikacije
urediU životinja i ljudi mikrosporidije često uzrokuju hronične, oslabljujuće bolesti a ne smrtonosne infekcije. Učinci na domaćina uključuju smanjenu dugovječnost, plodnost, težinu i opću energičnost. Često se izvještava o vertikalnom prijenosu mikrosporidija. U slučaju insekata domaćina, vertikalni prijenos često se događa transovarijski, tako da mikrosporidijski paraziti prelaze iz jajnika ženke domaćina u jajašce i na kraju se razmnožavaju u zaraženim larvama. Amblyospora salinaria n. sp. koji zarazi komarca Culex salinarius Coquillett i Amblyospora californica koji zarazi komarcem Culex tarsalis Coquillett, navode se kao tipski primjeri transovarijskog prijenosa mikrosporidija.[13][14][15][16]
Mikrosporidije, tačnije infekcije komaraca Vavraia culicis, istražuju se kao mogući obrazac razvoja mjera protiv malarij.[17] Mikrosporidijska infekcija Anopheles gambiae (glavni vektor malarije je Plasmodium falciparum) smanjuje malarijsku infekciju unutar komaraca i skraćuje im životni vijek.[18]
Kako većina komaraca zaraženih malarijom prirodno umire prije nego što je parazit malarije dovoljno zreo za prijenos, svako povećanje smrtnosti komaraca putem mikrosporidijanske infekcije može umanjiti prijenos malarije na ljude.
Klinčki značaj
urediMikrosporidijske infekcije ljudi ponekad uzrokuju bolest koja se zove mikrosporidioza. Najmanje 14 mikrosporidijanskih vrsta, iz osam rodova, prepoznate su kao ljudski patogeni. Oni uključuju Trachipleistophora hominis.[19]
Hiperparazitizam
uredi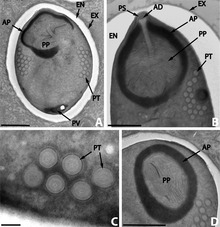
Mikrosporidije mogu zaraziti različite domaćine, uključujući i one koji su i sami paraziti. U tom slučaju, mikrosporidijska vrsta jeste hiperparazit, tj. parazit parazita. Kao primjer, poznato je više od osamnaest vrsta koje parazitiraju a Digenea (parazitske pljosnate gliste). Ove gliste su i same paraziti u raznim kičmenjacima i mehkušci ma. Osam ovih vrsta pripada rodu Nosema .[20]
Genomi
urediMikrosporidije imaju najmanje poznate (nuklearne) eukariotske genom e. Parazitski način života mikrosporidija doveo je do gubitka mnogih mitohondrija l i Golgijevih gena, pa čak su i njihove ribosomnne RNK manje u usporedbi s onima kod većine eukariota. Kao posljedica toga, genomi mikrosporidija su mnogo manji nego kod ostalih eukariota. ]]Do sada poznati mikrosporidijski genomi su veličine od 2,5 do 11,6 [[bazni par|megabaza, a kodiraju od 1.848 do 3.266 proteina što je u istom rasponu koliko i kod bakterija.[21] Izgleda da se u mikrosporidiji događalo više puta. Na primjer, genomi Encephalitozoon romaleae i Trachipleistophora hominis sadrže gene koji potiču od životinja i bakterija, a neki čak i iz gljiva. Izgleda da se kod mikrosporidija više puta dogodio horizontalni transfer gena (HTG). Naprimjer, genomi Encephalitozoon romaleae i Trachipleistophora hominis sadrže derivirane gene životinja i bakterija, a kod nekih, čak i gljiva.[21]
Klasifikacija
uredi

Mikrosporidije sa Aphelida i grupom Cryptomycota čine poseban takson.[22]
Prvi opisani rod mikrosporidija, Nosema, inicijalno je Nägeli stavio u skupinu gljiva Schizomycetes zajedno s nekim bakterijama i kvascima.[23][24] Neko vrijeme mikrosporidije su smatrane vrlo primitivnim eukariotima, smještenim u protozojsku skupinu Cnidospora.[3] Kasnije, posebno zbog nedostatka mitohondrija, premještene su zajedno s ostalim Protozoaama poput Diplomonada, Parabasalida i Archamoebae u protozojsku grupu Archezoa.[25] Novija istraživanja odbacila su ovu teorij ranog porijekla (za sve navedeno). Ipak, predlažu se da su mikrosporidije visoko razvijeni i specijalizovani organizmi, koji jednostavno više nemaju potrebne funkcije, jer ih opskrbljuje domaćin.[26] Nadalje, organizmi koji stvaraju spore uopće imaju složen sistem razmnožavanja, i seksualni i aseksualni, koji izgledaju daleko od primitivnih.
Danas su mikrosporidije smještene unutar gljiva ili kao sestrinska grupa gljiva sa zajedničkim predakom.[27][28][29][30]
Formiranje kladusa uglavnom se temelji na staništu i domaćinu. Vossbrinck i Debrunner-Vossbrinck, na osnovu staništa, predlažu tri klase mikrosporidija: Aquasporidia, Marinosporidia i Terresporidia.[31] Druga klasifikacija, mogla bi biti:[32]
- Razred Metchnikovellea Weiser 1977 emend. Cavalier-Smith 1993 [Manubrispora Cavalier-Smith 1998]
- Red Metchnikovellida Vivier 1977
- Porodica Amphiacanthidae
- Porodica Metchnikovellidae Caullery & Mesnil 1897
- Red Metchnikovellida Vivier 1977
- Razred Microsporea Delphy 1936 ex Levine & Corliss 1963
- Porodica Cougourdellidae Poisson 1953
- Porodica Facilisporidae
- Porodica Heterovesiculidae Lange et al. 1995
- Porodica Myosporidae
- Porodica Nadelsporidae
- Porodica Neonosemoidiidae
- Porodica Ordosporidae
- Porodica Pseudonosematidae
- Porodica Telomyxidae Leger & Hesse 1910
- Porodica Toxoglugeidae
- Porodica Tubulinosematidae
- Podrazred Haplophasea Sprague, Becnel & Hazard 1992
- Red Chytridiopsida Weiser 1974
- Porodica Chytridiopsidae Sprague, Ormieres & Manier 1972
- Porodica Buxtehudiidae Larsson 1980
- Porodica Enterocytozoonidae
- Porodica Burkeidae Sprague 1977
- Porodica Hesseidae Ormierees & Sprague 1973
- Red Glugeida Issi 1986
- Porodica Glugeidae Thelohan 1892 [Pleistophoridae Stempell 1909]
- Porodica Gurleyidae Sprague 1977
- Porodica Encephalitozoonidae
- Porodica Abelsporidae
- Porodica Tuzetiidae Sprague, Tuzet & Maurand 1977
- Porodica Microfilidae
- Porodica Unikaryonidae Sprague 1977
- Red Chytridiopsida Weiser 1974
- Podrazred Dihaplophasea Sprague, Becnel & Hazard 1992
- Red Meiodihaplophasida Sprague, Becnel & Hazard 1992
- Natporodica Thelohanioidea Hazard & Oldacre 1975
- Porodica Thelohaniidae Hazard & Oldacre 1975 [Coccosporidae]
- Porodica Duboscqiidae Sprague 1977
- Porodica Janacekiidae
- Porodica Pereziidae Loubes et al. 1977
- Porodica Striatosporidae
- Porodica Cylindrosporidae
- Natporodica Burenelloidea Jouvenaz & Hazard 1978
- Porodica Burenellidae Jouvenaz & Hazard 1978
- SuperPorodica Amblyosporoidea Weiser 1977
- Porodica Amblyosporidae Weiser 1977
- Natporodica Thelohanioidea Hazard & Oldacre 1975
- Red Dissociodihaplophasida Sprague, Becnel & Hazard 1992
- Natorodica Nosematoidea Labbe
- Porodica Nosematidae Labbe 1899
- Porodica Ichthyosporidiidae
- Porodica Caudosporidae Weiser 1958
- Porodica Pseudopleistophoridae Sprague 1977
- Porodica Mrazekiidae Leger & Hesse 1922
- Natorodica Culicosporoidea Weiser 1977
- Porodica Culicosporidae Weisser 1977
- Porodica Culicosporellidae
- Porodica Golbergiidae
- Porodica Spragueidae Weissenberg 1976
- Natporodica Ovavesiculoidea Sprague, Becnel & Hazard 1992
- Porodica Ovavesiculidae
- Porodica Tetramicridae
- Natorodica Nosematoidea Labbe
- Red Meiodihaplophasida Sprague, Becnel & Hazard 1992
Treća reklasifikacija jeCavalier-Smith 1993:[33]
- Potkoljeno Rudimicrospora Cavalier-Smith 1993
- Razred Minisporea Cavalier-Smith 1993
- Red Minisporida Sprague, 1972
- Razred Metchnikovellea Weiser, 1977
- Red Metchnikovellida Vivier, 1975
- Razred Minisporea Cavalier-Smith 1993
- Potkoljeno Polaroplasta Cavalier-Smith 1993
- Razred Pleistophoridea Cavalier-Smith 1993
- Red Pleistophorida Stempell 1906
- Razred Disporea Cavalier-Smith 1993
- Podrazred Unikaryotia Cavalier-Smith 1993
- Podrazred Diplokaryotia Cavalier-Smith 1993
- Razred Pleistophoridea Cavalier-Smith 1993
Također pogledajte
urediReference
uredi- ^ Delphy, J. 1936. Sous-règne des Protozoaires. In: Perrier, R. (ed.). La Faune de la France en tableaux synoptiques illustrés, v. 1A. Delagrave: Paris.
- ^ Levine, N. D. et al. (1980). A Newly Revised Classification of the Protozoa. The Journal of Protozoology 27: 37-58, [1].
- ^ a b Corliss JO, Levine ND (1963). "Establishment of the Microsporidea as a new class in the protozoan subphylum Cnidospora". The Journal of Protozoology. 10 (Suppl): 26–27. doi:10.1111/jeu.1963.10.issue-s3.
- ^ Sprague, V. (1977). Classification and phylogeny of the Microsporidia. In: Comparative pathobiology. vol. 2, Systematics of the Microsporidia. Lee A. Bulla & Thomas C. Cheng (ed.). pp. 1-30. New York: Plenum Press, [2].
- ^ Hibbett, D.S. et al. (2007) A higher level phylogenetic classification of the Fungi. Mycological Research 111 (5): 509–47, [3].
- ^ Silar, Philippe (2016). Protistes Eucaryotes : Origine, Evolution et Biologie des Microbes Eucaryotes. HAL. str. 462. ISBN 978-2-9555841-0-1.
- ^ Hawksworth, David (2001). "The magnitude of fungal diversity: The 1.5 million spices estimate revisited". Mycological Research. 105 (12): 1422. doi:10.1017/S0953756201004725.
- ^ a b Ronny Larsson, Lund University (Department of Cell and Organism Biology) Cytology and taxonomy of the microsporidia Arhivirano 12. 9. 2009. na Wayback Machine 2004.
- ^ Didier, ES. (Apr 2005). "Microsporidiosis: an emerging and opportunistic infection in humans and animals". Acta Trop. 94 (1): 61–76. doi:10.1016/j.actatropica.2005.01.010. PMID 15777637.
- ^ Hoffman, G. (1999). Parasites of North American Freshwater Fishes, 2nd edn, University of California Press, Berkeley, California, USA, p. 89, [4].
- ^ Winters, A. D., Faisal, M. 2014. Molecular and ultrastructural characterization of Dictyocoela diporeiae n. sp. (Microsporidia), a parasite of Diporeia spp. (Amphipoda, Gammaridea). Parasite, 21, 26 doi:10.1051/parasite/2014028

- ^ Ironside JE (2007). "Multiple losses of sex within a single genus of Microsporidia". BMC Evolutionary Biology. 7: 48. doi:10.1186/1471-2148-7-48. PMC 1853083. PMID 17394631.
- ^ Andreadis TG, Hall DW (august 1979). "Development, ultrastructure, and mode of transmission of Amblyospora sp. (Microspora) in the mosquito". The Journal of Protozoology. 26 (3): 444–52. doi:10.1111/j.1550-7408.1979.tb04651.x. PMID 536933.
- ^ Andreadis TG, Hall DW (septembar 1979). "Significance of transovarial infections of Amblyospora sp. (Microspora:Thelohaniidae) in relation to parasite maintenance in the mosquito Culex salinarius". Journal of Invertebrate Pathology. 34 (2): 152–7. doi:10.1016/0022-2011(79)90095-8. PMID 536610.
- ^ Jahn GC, Hall DW, Zam SG (1986). "A comparison of the life cycles of two Amblyospora (Microspora: Amblyosporidae) in the mosquitoes Culex salinarius and Culex tarsalis Coquillett". Journal of the Florida Anti-Mosquito Association. 57 (1): 24–27.
- ^ Becnel JJ, Andreadis TG (maj 1998). "Amblyospora salinaria n. sp. (Microsporidia: amblyosporidae), parasite of Culex salinarius (Diptera: culicidae): its life cycle stages in an intermediate host". Journal of Invertebrate Pathology. 71 (3): 258–62. doi:10.1006/jipa.1998.4729. PMID 9538031.
- ^ Koella, Jacob C.; Lorenz, Lena; Bargielowski, Irka (2009). Chapter 12 Microsporidians as Evolution-Proof Agents of Malaria Control?. Advances in Parasitology. 68. str. 315–327. doi:10.1016/S0065-308X(08)00612-X. ISBN 978-0-12-374787-7. PMID 19289199.
- ^ Bargielowski I, Koella JC (2009). Baylis M (ured.). "A Possible Mechanism for the Suppression of Plasmodium berghei Development in the Mosquito Anopheles gambiae by the Microsporidian Vavraia culicis". PLoS ONE. 4 (3): e4676. Bibcode:2009PLoSO...4.4676B. doi:10.1371/journal.pone.0004676. PMC 2651578. PMID 19277119.
- ^ Heinz, E; Williams, TA; Nakjang, S; et al. (Oct 2012). "The genome of the obligate intracellular parasite Trachipleistophora hominis: New insights into microsporidian genome dynamics and reductive evolution". PLoS Pathog. 8 (10): e1002979. doi:10.1371/journal.ppat.1002979. PMC 3486916. PMID 23133373.
- ^ a b Toguebaye, B. S.; Quilichini, Y.; Diagne, P. M.; Marchand, B. (2014). "Ultrastructure and development of Nosema podocotyloidis n. sp. (Microsporidia), a hyperparasite of Podocotyloides magnatestis (Trematoda), a parasite of Parapristipoma octolineatum (Teleostei)". Parasite. 21: 44. doi:10.1051/parasite/2014044. PMC 4150386. PMID 25174849.
- ^ a b Corradi, N.; Selman, M. (2013). "Latest Progress in Microsporidian Genome Research". Journal of Eukaryotic Microbiology. 60 (3): 309–312. doi:10.1111/jeu.12030. PMID 23445243.
- ^ Karpov Sergey, Mamkaeva Maria A, Aleoshin Vladimir, Nassonova Elena, Lilje Osu, Gleason Frank H. Morphology, phylogeny and ecology of the aphelids (Aphelidea, Opisthokonta) and proposal for the new superphylum Opisthosporidia. 5. Frontiers in Microbiologyyear= 2014. doi:10.3389/fmicb.2014.00112.CS1 održavanje: više imena: authors list (link)
- ^ Nägeli, C. von (1857). Über die neue Krankheit der Seidenraupe und verwandte Organismen. pp. 760–61. In: Caspary, R. (ed.). Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten in Bonn von 18 bis 24 September 1857. Botanische Zeitung 15: 749–776, [5].
- ^ Keeling, P. J., & Fast, N. M. (2002). Microsporidia: biology and evolution of highly reduced intracellular parasites. Annual Reviews in Microbiology, 56(1), 93-116, [6].
- ^ Cavalier-Smith, T. (1993). Kingdom protozoa and its 18 phyla. Microbiological Reviews 57 (4): 953–994, [7].
- ^ Keeling PJ, Slamovits CH (decembar 2004). "Simplicity and Complexity of Microsporidian Genomes". Eukaryotic Cell. 3 (6): 1363–9. doi:10.1128/EC.3.6.1363-1369.2004. PMC 539024. PMID 15590811.
- ^ Fischer WM, Palmer JD (septembar 2005). "Evidence from small-subunit ribosomal RNA sequences for a fungal origin of Microsporidia". Molecular Phylogenetics and Evolution. 36 (3): 606–22. doi:10.1016/j.ympev.2005.03.031. PMID 15923129.
- ^ Liu YJ, Hodson MC, Hall BD (2006). "Loss of the flagellum happened only once in the fungal lineage: phylogenetic structure of Kingdom Fungi inferred from RNA polymerase II subunit genes". BMC Evolutionary Biology. 6: 74. doi:10.1186/1471-2148-6-74. PMC 1599754. PMID 17010206.
- ^ Gill EE, Fast NM (juni 2006). "Assessing the microsporidia-fungi relationship: Combined phylogenetic analysis of eight genes". Gene. 375: 103–9. doi:10.1016/j.gene.2006.02.023. PMID 16626896.
- ^ Lee SC, Corradi N, Byrnes EJ, et al. (novembar 2008). "Microsporidia evolved from ancestral sexual fungi". Current Biology. 18 (21): 1675–9. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2008.09.030. PMC 2654606. PMID 18976912.
- ^ Vossbrinck CR, Debrunner-Vossbrinck BA (maj 2005). "Molecular phylogeny of the Microsporidia: ecological, ultrastructural and taxonomic considerations". Folia Parasitologica. 52 (1–2): 131–42, discussion 130. doi:10.14411/fp.2005.017. PMID 16004372.
- ^ Alimov, A. F., ured. (maj 2007). "Protista 2: Handbook on zoology". Nauka: 1141. journal zahtijeva
|journal=(pomoć) - ^ Cavalier-Smith (1993). "Kingdom Protozoa and its 18 phyla" (PDF). journal zahtijeva
|journal=(pomoć)
Vanjski linkovi
uredi- BioHealthBase Bioinformatics Resource Center Database of microspordia sequences and related information.
- Microsporidia na US National Library of Medicine Medical Subject Headings (MeSH)