Senzorni neuron
Senzorni neuroni ili čulni neuroni, takođe poznati kao aferentni neuroni, su neuroni u nervnom sistemu, koji pretvaraju određeni tip stimulusa, preko njihovih receptora, u akcijski potencijal ili gradirani potencijal.[1] Ovaj proces zove se senzorna transdukcija. Ćelijska tijela senzornih neurona nalaze se u dorzalnoj gangliji kičmene moždine.[2]
_E.PNG)
Senzorne informacije putuju preko aferentnih nervnih vlakana u čulnom nervu, do mozga preko kičmene moždine. Podražaj može doći od "eksteroreceptora" izvan tijela, naprimjer onih koji detektuju svjetlost i zvuk, ili od "interoreceptora" unutar tijela, naprimjer onih koji reaguju na krvni pritisak ili osećaj položaja tijela.
Tipovi i funkcije
urediRazličiti tipovi senzornih neurona imaju različite senzorne receptore koji reaguju na različite tipove podražaja. Postoji najmanje šest vanjskih i dva unutrašnja senzorna receptora:
Vanjski receptori
urediVanjski receptori koji reaguju na podražaje izvan tijela nazivaju se eksteroreceptori.[3] Eksteroreceptori uključuju olfaktorne receptore (miris), receptore okusa, fotoreceptore (vid), trapljaste ćelije (sluh), termoreceptore ( temperatura), i niz različitih mehanoreceptora (rastezanje, izobličenje)
Miris
urediSenzorni neuroni uključeni u miris nazivaju se senzorni neuroni njuha. Ovi neuroni sadrže receptore, zvane olfaktorni receptori, koje aktiviraju mirisne molekule u zraku. Molekule u zraku detektuju se uvećanim trepljama] i mikroresicama.[4] Ovi senzorni neuroni proizvode akcijske potencijale. Njihovi akson[]i formiraju olfaktorni nerv i sinapsiraju direktno na neurone u moždanoj kori (olfaktornu bulbus). Oni ne koriste isti put kao drugi senzorni sistemi, zaobilazeći moždano stablo i talamus. Neuroni u olfaktornom režnju koji primaju direktan senzorni nervni ulaz, imaju veze s drugim dijelovima olfaktornog sistema i mnogim dijelovima limbnog sistema.
Okus
urediSlično kao olfaktorni receptor, receptor pkusa u okusnom pupoljku stupaju u interakciju sa hemikalijama u hrani da bi proizveli akcijski potencijal.
Vid
urediFotoreceptorske ćelije su sposobne za fototransdukciju, proces koji pretvara svjetlost (elektromagnetno zračenje) u električne signale. Ovi signali se rafiniraju i kontroliraju interakcijama s drugim tipovima neurona u mrežnjačama. Pet osnovnih klasa neurona unutar retine su fotoreceptorske ćelije, bipolarne ćelije, ganglijske ćelije, horizontalne ćelije i amakrine ćelije . Osnovno kolo mrežnjače uključuje lanac od tri neurona koji se sastoji od fotoreceptora (bilo štapića ili čepića), bipolarne ćelije i ganglijske ćelije. Prvi akcijski potencijal javlja se u ganglijskoj ćeliji mrežnjače. Ovaj put je najdirektniji način za prenošenje vizualnih informacija u mozak. Postoje tri primarna tipa fotoreceptora: čepići (čunjići) su fotoreceptori koji značajno reaguju na boju. Kod ljudi tri različita tipa čunjića odgovaraju primarnom odgovoru na kratku talasnu dužinu (plava), srednju talasnu dužinu (zelena) i dugu talasnu dužinu (žuta/crvena).[5] Štapići su fotoreceptori koji su vrlo osjetljivi na intenzitet svjetlosti, omogućavajući vid pri slabom svjetlu. Koncentracija i omjer štapića i čunjića u snažnoj je korelaciji s tim da li je životinja dnevna ili noćna. Kod ljudi, štapići su brojniji od čunjića za otprilike 20:1, dok je kod noćnih životinja, kao što je sova, taj odnos bliži 1000:1.[5] Ćelije mrežnjačnog ganglijakog soja uključene su u simpatički odgovor. Od oko 1,3 miliona ganglijskih ćelija prisutnih u retini, veruje se da je 1-2% fotosenzitivnih.[6]
Problemi i propadanje senzornih neurona povezanih s vidom dovode do poremećaja kao što su:
- Makulska degeneracija – degeneracija centralnog vidnog polja zbog ćedlijskih ostataka ili krvnih žila koji se nakupljaju između mrežnjača i sudovnjače, čime se ometa i/ili uništava složena interakcija neurona koji su tamo prisutni.[7]
- Glaukom – gubitak ganglijskih ćelija mrežnjače što uzrokuje gubitak vida do sljepila.[8]
- Dijabeteska retinopatija – loša kontrola šećera u krvi zbog dijabetesa oštećuje sitne krvne sudove u mrežnjači.[9]
Sluh
urediSlušni sistem je odgovoran za pretvaranje talasa pritiska generisanih vibrirajućim zračnim molekulama ili zvukom u signale koje mozak može interpretirati.
Ova mehanoelektrična transdukcija je posredovana trepljasim ćelijama unutar uha. Ovisno o kretanju, trepljasta ćelija može ili hiperpolarizirati ili depolarizirati. Kada je kretanje prema najvišoj stereociliji, Na+ kationski kanali se otvaraju dozvoljavajući Na+ da teče u ćeliju i rezultirajuća depolarizacija uzrokuje Ca ++ kanali da se otvore, oslobađajući tako svoj neurotransmiter u aferentnim slušnim nervima. Postoje dva tipa trepljasrtih ćelija: unutrašnje i vanjske. Unutrašnje su senzorni receptori.[10]
Problemi sa senzornim neuronima povezanim sa slušnim sistemom dovode do poremećaja kao što su:
- Poremećaj slušne obrade – Auditorne informacije u mozgu se obrađuju na abnormalan način. Pacijenti s poremećajem slušne obrade obično mogu normalno dobiti informacije, ali njihov mozak ih ne može pravilno obraditi, što dovodi do oštećenja sluha.[11]
- Auditivna verbalna agnozija – Razumijevanje govora je izgubljeno, ali se zadržava sposobnost sluha, govora, čitanja i pisanja. Ovo je uzrokovano oštećenjem stražnjeg gornjeg sljepoočnih režnjeva režnjeva, što opet ne dozvoljava mozgu da pravilno obrađuje zvučni unos.[12]
Temperatura
urediTermoreceptori su senzorni receptori, koji reaguju na različite temperature. Dok su mehanizmi preko kojih ovi receptori djeluju nisu jasni, nedavna otkrića su pokazala da sisari imaju najmanje dva različita tipa termoreceptora.[13] Bulboidno tjelešce je kožni receptor "osjetljiv na hladnoću", koji detektuje niske temperature. Drugi tip je receptor osjetljiv na toplinu.
Mehanoreceptori
urediMehanoreceptori su senzorni receptori koji reaguju na mehaničke sile, kao što su pritisak ili izobličenje.[14]
Specijalizovane senzorne receptorske ćelije koje se nazivaju mehanoreceptori često inkapsuliraju aferentna vlakna, kako bi pomogle da se aferentna vlakna podese na različite tipove somatske stimulacije. Mehanoreceptori također pomažu u snižavanju pragova za stvaranje akcijsog potencijala u aferentnim vlaknima i na taj način povećavaju vjerovatnoću nadražaja u prisustvu senzorne stimulacije.[15]
Neki tipovi mehanoreceptora ispaljuju akcijske potencijale kada su njihove membrane fizički istegnute.
Proprioceptori su još jedan tip mehanoreceptora što doslovno znači "receptori za sebe". Ovi receptori pružaju prostorne informacije o udovima i drugim dijelovima tijela.[16]
Nociceptori su odgovorni za obradu bola i promjena temperature. Pekuća bol i iritacija nakon jedenja čili papričice (zbog njenog glavnog sastojka, kapsaicina), osjećaj hladnoće nakon uzimanja hemikalija kao što je mentol ili icilin, kao i uobičajeni osjećaj bola rezultat su neurona sa ovih receptora.[17]
Problemi sa mehanoreceptorima dovode do poremećaja kao što su:
- Neuropatski bol - stanje teške boli koje je rezultat oštećenog senzornog živca [17]
- Hiperalgezija - povećana osjetljivost na bol uzrokovana senzornim ionskim kanalom, TRPM8, koji tipski reaguje na temperature između 23 i 26 stepeni, i pruža osjećaj hlađenja povezan s mentolom i icilinom [17]
- Sindrom fantomskih udova - poremećaj senzornog sistema gdje se bol ili pokret doživljavaju u udu koji ne postoji [18]
Unutrašnji receptori
urediUnutrašnji receptori koji reaguju na promjene unutar tijela poznati su kao interoceptori.[3]
Krv
urediAortna tela i karotidna tela sadrže klastere glomusnih ćelija - perifernih hemoreceptora koji detektuju promjene hemijskih svojstava u krvi kao što su koncentracije kisika.[19] Ovi receptori polimodalno reaguju na brojne različite stimuluse.
Nociceptori
urediNociceptori reagiraju na potencijalno štetne podražaje slanjem signala kičmenoj moždini i mozgu. Ovaj proces, nazvan nocicepcija, obično uzrokuje percepciju bola.[20][21] Nalaze se u unutrašnjim organima kao i na površini tijela kako bi "otkrili i zaštitili".[21] Nociceptori otkrivaju različite tipove štetnih podražaja koji ukazuju na mogućnost oštećenja, a zatim pokreću nervne odgovore da se povuku iz stimulusa .[21]
- Termalni nociceptori se aktiviraju štetnom toplotom ili hladnoćom na različitim temperaturama.[21]
- Mehanički nociceptori reaguju na višak pritiska ili mehaničku deformaciju, kao što je štipanje.[21]
- „Hemijski“ nociceptori reaguju na širok spektar hemikalija, od kojih neke signaliziraju odgovor. Oni su uključeni u otkrivanje nekih začina u hrani, kao što su oštri sastojci u biljkama Brassica i Allium, koji ciljaju senzorne neuronske receptore da izazovu akutni bol i naknadnu preosjetljivost na bol.[22]
Veza sa centralnim nervnim sistemom
urediInformacije koje dolaze iz senzornih neurona u glavi ulaze u centralni nervni sistem (CNS) preko kranijalnih nerava. Informacije iz senzornih neurona ispod glave ulaze u kičmenu moždinu i prolaze prema mozgu kroz 31 kičmeni nerv.[23] Senzorne informacije koje putuju kroz kičmenu moždinu prate dobro definisane puteve. Nervni sistem kodira razlike među senzacijama u smislu toga koje ćelije su aktivne.
Klasifikacija
urediAdekvatan stimulans
urediAdekvati podražaj senzornog receptora je modalitet stimulacije za koji posjeduje adekvatan senzorna transdukcija aparata. Adekvatan stimulans se može koristiti za klasifikaciju senzornih receptora:
- Baroreceptori reaguju na pritisak u krvnim sudovima
- Hemoreceptori reaguju na hemijske stimuluse
- Receptori elektromagnetnog zračenja reaguju na elektromagnetno zračenje[24]
- Infracrveni receptori reaguju na infracrveno zračenje
- Fotoreceptori reaguju na vidljivo svjetlo
- Ultraljubičasti receptor reaguju na ultraljubičasto zračenje
- Electroreceptori reaguju na električno polje
- Lorenzinijeve ampule reaguju na električna polja, salinitet i temperaturu, ali funkcionišu prvenstveno kao elektroreceptori
- Hidroreceptor reaguju na promjene vlažnosti
- Magnetorceptori reaguju na magnetno polje
- Mehanoreceptori reaguju na mehaničko naprezanje ili mehaničko istezanje
- Nociceptori reaguju na oštećenje ili prijetnju oštećenja tjelesnih tkiva, što dovodi (često, ali ne uvijek) do percepcije bola
- Osmoreceptori reaguju na osmolarnost tečnosti (kao što je hipotonija)
- Proprioceptori pružaju osećaj položaja
- Termoreceptori reaguju na temperaturu, bilo toplotu, hladnoću ili oboje
Lokacija
urediSenzorni receptori se mogu klasificirati prema lokaciji:
- Kožni receptori su senzorni receptori koji se nalaze u dermi ili epidermi.[25]
- Mišićno vreteno sadrže mehanoreceptore koji detektuju istezanje mišića.
Morfologija
urediSomatski senzorni receptori blizu površine kože se obično mogu podijeliti u dvije grupe na osnovu morfologije:
- Slobodni nervni završeci karakteriziraju nociceptore i termoreceptore i nazivaju se tako jer su terminalne grane neurona nemijelinizirane i šire se po dermisu i epidermi.
- Inkapsulirani receptor se sastoje od preostalih tipova kožnih receptora. Enkapsulacija postoji za specijalizirano funkcioniranje.
Stopa adaptacije
uredi- Tonijski receptor je senzorni receptor koji se polako prilagođava na stimulus [26] and continues to produce action potentials over the duration of the stimulus.[27] Na taj način prenosi informaciju o trajanju stimulusa. Neki tonijski receptori su trajno aktivni i ukazuju na pozadinski nivo. Primjeri takvih tonijskih receptora su receptori bola, zglobna kapsula i mišićno vreteno.[28]
- Fazični receptor je senzorni receptor koji se brzo prilagođava stimulusu. Reakcija ćelije se vrlo brzo smanjuje, a zatim prestaje.[29] Ne daje informacije o trajanju stimulusa;[27] umjesto toga neki od njih prenose informacije o brzim promjenama intenziteta i brzine stimulusa.[28] Primjer fazni receptor je Pacinijevo telo.
Lijekovi
urediTrenutno na tržištu postoji mnogo lijekova koji se koriste za manipulaciju ili liječenje poremećaja senzornog sistema. Naprimjer, Gabapentin je lijek koji se koristi za liječenje neuropatskog bola interakcijom s jednim od naponsko zavisnih kalcijskih kanala prisutnih na nereceptivnim neuronima.[17] Neki lijekovi mogu biti koristi se za borbu protiv drugih zdravstvenih problema, ali može imati neželjene nuspojave na senzorni sistem. Ototoksični lijekovi su lijekovi koji utiču na pužnicu upotrebom toksina poput aminoglikozidnih antibiotika, koji truju ćelije kose. Korištenjem ovih toksina, trepljaste ćelije koje pumpaju K+ prestaju funkcionirati. Tako se gubi energija koju generiše endopužnični potencijal koji pokreće proces transdukcije slušnog signala, što dovodi do gubitka sluha.[30]
Neuroplastičnost
urediOtkako su naučnici primetili korteksno remapiranje u mozgu Tauba Silver Spring majmuna, postoji velika količina istraživanja plastičnosti senzornog sistema. Ogroman napredak je napravljen u liječenju poremećaja senzornog sistema. Tehnike kao što je terapija pokreta izazvana ograničenjem koju je razvio Taub pomogle su pacijentima s paraliziranim udovima da ponovo koriste svoje udove prisiljavajući senzorni sistem da razvija nove nervne puteve.[31] Sindrom fantomskih udova je poremećaj senzornog sistema u kojem amputirci percipiraju da im amputirani ud još uvijek postoji i da još uvijek mogu osjećati bol u njemu. ogledalska kutija razvijena od strane V.S. Ramachandrana, omogućio je pacijentima sa sindromom fantomskih udova da ublaže percepciju paraliziranih ili bolnih fantomskih udova. To je jednostavan uređaj koji koristi ogledalo u kutiji kako bi stvorio iluziju u kojoj senzorni sistem percipira da vidi dvije ruke umjesto jedne, što omogućava senzornom sistemu da kontroliše "fantomski ud". Na taj način se senzorni sistem može postepeno priviknuti na amputirani ekstremitet i tako ublažiti ovaj sindrom.[32]
Ostale životinje
urediHidrodinamička recepcija je oblik mehanorecepcije koji se koristi u nizu životinjskih vrsta.
Dodatne slike
uredi-
 Ilustracija taktilnih receptora u koži
Ilustracija taktilnih receptora u koži -
 Ilustracija lameliranog tjelašceta
Ilustracija lameliranog tjelašceta -
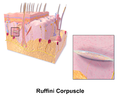 Ilustracija Ruffinijevog tijela
Ilustracija Ruffinijevog tijela -
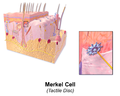 Ilustracija kožne Merkelove ćelije
Ilustracija kožne Merkelove ćelije -
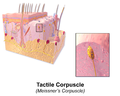 Ilustracija taktilnog tjelašceta
Ilustracija taktilnog tjelašceta -
 Ilustracija pleksusa korijena dlake
Ilustracija pleksusa korijena dlake -
 Ilustracija slobodnih nervnih završetaka
Ilustracija slobodnih nervnih završetaka







Također pogledajte
urediReference
uredi- ^ Parsons, Richard (2018). CGP: A-Level Biology Complete Revision & Practice. Newcastle Upon Thynde: Coordination Group Publishing Ltd. str. 138. ISBN 9781789080261.
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 izd.). Sinauer Associates, Inc. str. 207. ISBN 978-0878936977.
- ^ a b Campbell, Neil (1996). Biology (4th izd.). Benjamin/Cummings Pub. Co. str. 1028. ISBN 0805319409.
- ^ Breed, Michael D., and Moore, Janice. Encyclopedia of Animal Behavior . London: Elsevier, 2010. Print.
- ^ a b "eye, human." Encyclopædia Britannica. Encyclopædia Britannica Ultimate Reference Suite. Chicago: Encyclopædia Britannica, 2010.
- ^ Foster, R. G.; Provencio, I.; Hudson, D.; Fiske, S.; Grip, W.; Menaker, M. (1991). "Circadian photoreception in the retinally degenerate mouse (rd/rd)". Journal of Comparative Physiology A 169. doi:10.1007/BF00198171
- ^ de Jong, Paulus T.V.M. (5. 10. 2006). "Age-Related Macular Degeneration". New England Journal of Medicine. 355 (14): 1474–1485. doi:10.1056/NEJMra062326. ISSN 0028-4793. PMID 17021323.
- ^ Alguire, Patrick; Dallas, Wilbur; Willis, John; Kenneth, Henry (1990). "Chapter 118 Tonometry". Clinical methods : the history, physical, and laboratory examinations (3 izd.). Butterworths. ISBN 978-0409900774. OCLC 15695765.
- ^ "NIHSeniorHealth: Diabetic Retinopathy - Causes and Risk Factors". nihseniorhealth.gov. Arhivirano s originala, 14. 1. 2017. Pristupljeno 19. 12. 2016.
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 izd.). Sinauer Associates, Inc. str. 327–330. ISBN 978-0878936977.
- ^ "Auditory Processing Disorder (APD)" (PDF). British Society of Audiology APD Special Interest Group MRC Institute of Hearing Research. Arhivirano s originala (PDF), 2. 4. 2016. Pristupljeno 22. 4. 2023.
- ^ Stefanatos, Gerry A.; Gershkoff, Arthur; Madigan, Sean (1. 7. 2005). "On pure word deafness, temporal processing, and the left hemisphere". Journal of the International Neuropsychological Society. 11 (4): 456–470, discussion 455. doi:10.1017/S1355617705050538. ISSN 1355-6177. PMID 16209426. S2CID 25584363.
- ^ Krantz, John. Experiencing Sensation and Perception Arhivirano 17. 11. 2017. na Wayback Machine. Pearson Education, Limited, 2009. p. 12.3
- ^ Winter, R., Harrar, V., Gozdzik, M., & Harris, L. R. (2008). The relative timing of active and passive touch. [Proceedings Paper]. Brain Research, 1242, 54-58. doi:10.1016/j.brainres.2008.06.090
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 izd.). Sinauer Associates, Inc. str. 209. ISBN 978-0878936977.
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 izd.). Sinauer Associates, Inc. str. 215–216. ISBN 978-0878936977.
- ^ a b c d Lee, Y; Lee, C; Oh, U (2005). "Painful channels in sensory neurons". Molecules and Cells. 20 (3): 315–324. PMID 16404144.
- ^ Halligan, Peter W; Zeman, Adam; Berger, Abi (4. 9. 1999). "Phantoms in the brain". BMJ: British Medical Journal. 319 (7210): 587–588. doi:10.1136/bmj.319.7210.587. ISSN 0959-8138. PMC 1116476. PMID 10473458.
- ^ Satir, P. & Christensen, S.T. (2008) Structure and function of mammalian cilia. in Histochemistry and Cell Biology, Vol 129:6
- ^ Sherrington C. The Integrative Action of the Nervous System. Oxford: Oxford University Press; 1906.
- ^ a b c d e St. John Smith, Ewan (14. 10. 2017). "Advances in understanding nociception and neuropathic pain". Journal of Neurology. 265 (2): 231–238. doi:10.1007/s00415-017-8641-6. ISSN 0340-5354. PMC 5808094. PMID 29032407.
- ^ Zhao, Jianhua; Lin King, John V.; Paulsen, Candice E.; Cheng, Yifan; Julius, David (8. 7. 2020). "Irritant-evoked activation and calcium modulation of the TRPA1 receptor". Nature. 585 (7823): 141–145. Bibcode:2020Natur.585..141Z. doi:10.1038/s41586-020-2480-9. ISSN 1476-4687. PMC 7483980. PMID 32641835.
- ^ Kalat, James W. (2013). Biological Psychology (11th izd.). Wadsworth Publishing. ISBN 978-1111831004.
- ^ Michael J. Gregory. "Sensory Systems". Clinton Community College. Arhivirano s originala, 25. 6. 2013. Pristupljeno 6. 6. 2013.
- ^ "Cutaneous receptor".
- ^ Binder, Marc D.; Hirokawa, Nobutaka; Windhorst, Uwe (2009). Encyclopedia of neuroscience ([Online-Ausg.]. izd.). Berlin: Springer. ISBN 978-3-540-29678-2.
- ^ a b mentor.lscf.ucsb.edu/course/fall/eemb157/lecture/Lectures%2016,%2017%2018.ppt [mrtav link]
- ^ a b "Sensory Receptor Function". frank.mtsu.edu. Arhivirano s originala, 3. 8. 2008.
- ^ Sherwood, Lauralee; Klandorf, Hillar; Yancey, Paul (2012). Animal Physiology: From Genes to Organisms (jezik: engleski). Cengage Learning. ISBN 978-0840068651. Pristupljeno 13. 12. 2017.
- ^ Priuska, E.M.; Schacht, J. (1997). "Mechanism and prevention of aminoglycoside ototoxicity: Outer hair cells as targets and tools". Ear, Nose, & Throat Journal. 76 (3): 164–171. doi:10.1177/014556139707600310. PMID 9086645. S2CID 8216716.
- ^ Schwartz and Begley 2002, p. 160; "Constraint-Induced Movement Therapy", excerpted from "A Rehab Revolution," Stroke Connection Magazine, September/October 2004. Print.
- ^ Blakeslee, Sandra; Ramachandran, V. S. (1998). Phantoms in the brain : probing the mysteries of the human mind. William Morrow & Company, Inc. ISBN 978-0688152475. OCLC 43344396.