Riblja krljušt
Riblja krljušt je mala kruta ploča koja raste iz kože riba. Koža većine riba s vilicama prekrivena je ovim zaštitnim ljuskama, koje također mogu pružiti efikasnu kamuflažu korištenjem refleksije i boja, kao i moguće hidrodinamičke prednosti. Termin krljušt potiče od starofrancuskog escale, što znači mahuna ili ljuska.[1]

Krljušti se enormno razlikuju po veličini, obliku, strukturi i opsegu, u rasponu od jakih i čvrstih oklopnih ploča kod riba kao što su škampaste ribe i tropske morske ribe sa krljuštima od koštanih ploča koje okružuju tijelo, iz kojih strše bodlje, do mikroskopskih ili ih nema kod riba kao što su jegulja i morske ribe. Morfologija krljušti može se koristiti za identifikaciju vrste ribe od koje potiče. Krljušti su nastale unutar Ostracoderma bez vilica, predaka svih današnjih riba s čeljustima. Većina koštanih riba prekrivena je cikloidnim ljuskama lososa i šarana, ili ktenoidnim ljuskama smuđa, ili ganoidnim ljuskama jesetra i garovi. Hrskavicees (ajkules izrake) su prekrivene plakoidnim ljuskama. Neke vrste su umjesto toga prekrivene scutes, a druge nemaju vanjski omotač na dijelu ili cijeloj koži.
Riblje krljušti su dio ribljeg pokrovnog sistema, a proizvode se od mezodermskog sloja dermisa, što ih razlikuje od gmizavačkih ljuski.[2][3] Isti geni uključeni u razvoj zuba i dlake kod sisara su također uključeni u razvoj krljušti. Plakoidne krljušti hrskavičavih riba nazivaju se i dermski zubići i strukturno su homologne sa zubima kičmenjaci|kičmenjaka]]. Većina riba je također prekrivena slojem sluzi ili materijom sličnom sluzi koja može zaštititi od patogena kao što su bakterije, gljive i virusi, i smanjiti površinski otpor kada riba pliva.
Telodontne krljušti
uredi
Koštane krljušti thelodonta, najzastupljenijeg oblika fosilnih riba, dobro su poznate. Formirale su se i raspadale tokom života organizama, i brzo su se razdvajale nakon njihove smrti.[4]
Kost, tkivo koje je i otporno na mehanička oštećenja i relativno sklono fosilizaciji, često čuva unutrašnje detalje, što omogućava detaljno proučavanje histologije i rasta krljušti. Sastoje sed od nerastuće "krune" sastavljene od dentina, sa ponekad ukrašenom gornjom površinom cakline i aspidinskom bazom.[5] Njihova rastuća osnova je napravljena od kosti bez ćelija, koja je ponekad razvila strukture sidrišta kako bi je učvrstila u boku ribe.[6] Osim toga, čini se da postoji pet tipova rasta kosti, koji mogu predstavljati pet prirodnih grupa unutar telodonta – ili spektar koji se kreće između krajnjih članova meta- (ili orto-) dentinskog i mezodentinskog tkiva.[7] Čini se da svaki od pet oblika liči na krljušti izvedenijih grupa riba, što sugerira da su telodontske grupe možda bile grupe stabala narednih skupina riba.[6]
Međutim, korištenje samo morfologije krljušti za razlikovanje vrsta ima neke zamke. Unutar svakog organizma, oblik krljušti uveliko varira ovisno o površini tijela,[8] sa intermedijarnim oblicima koji se pojavljuju između različitih područja - i da stvar bude gora, morfologija krljušti možda čak nije ni konstantna unutar jednog područja. Da stvari dodatno zbune, morfologije im nisu jedinstvene za taksone i mogu se ne razlikovati na istom području dvije različite vrste.[9]
Morfologija i histologija telodonta predstavljaju glavni alat za kvantificiranje njihove raznolikosti i razlikovanje vrsta, iako je korištenje takvih konvergentnih osobina sklono greškama. Ipak, predložen je okvir koji se sastoji od tri grupe, na osnovu morfologije i histologije krljušti.[7] Poređenja s modernim vrstama morskih pasa pokazala su da su krljušti telodonta bile funkcionalno slične onima modernih hrskavičnih riba, a također su omogućile opsežnu usporedbu između ekoloških niša.[10]
Kosmoide krljušti
uredi
Kosmoidne krljušti nalaze se samo na drevnim riba s perajima, uključujući neke od najranijih plučašica (podrazred Dipnoi), i u Crossopterygii, uključujući današnje Coelacantha u modificiranom obliku (vidi elazmoidne ljuske, dolje). Vjerovatno su izvedeni iz fuzije plakoidno-ganoidnih ljuskica. Unutrašnji dio krljušti napravljen je od guste lamelirane kosti zvane izopedin. Povrh toga leži sloj spužvaste ili vaskularne kosti opskrbljene krvnim sudovima, nakon čega slijedi složeni sloj nalik dentinu koji se zove kosmin, s površinskom vanjskom prevlakom od vitrodentina. Gornja površina je keratin. Kosmoidne ljuske povećavaju se u veličini rastom sloja lamelirane kosti.[11]
Elazmoidne krljušti
uredi
Elazmoidne krljušti su tanke, imbricirane ljuske sastavljene od sloja guste, lamelirane kolagenske kosti zvane izopedin, iznad koje se nalazi sloj tuberkula obično sastavljen od kosti, kao u rodu Eusthenopteron. Sloj dentina koji je bio prisutan u ribama s prvim režnjevitimm perajima obično je smanjen, kao kod postojećeg Coelacantha, ili potpuno odsutan, kao kod postojećih dvodihalica i u devonskom rodu Eusthenopteron.[12] Elazmoidne krljušti su se pojavile nekoliko puta tokom evolucije riba. Prisutni su u nekim ribama s režnjevim perajima, kao što su sve postojeće i neke izumrle plućašice, kao i Coelacanthi koje imaju modificiranu kosmoidnu ljusku kojoj nedostaje kosmin i tanje su nego prave kosmoidne krljušti. Prisutni su i kod nekih tetrapodomorfa poput Eusthenopteron, amiida i teleostea, čije cikloidne i ktenoidne krljušti predstavljaju najmanje mineralizirane elazmoidne krljušti.
Elazmoidne krljušti zebrica koriste se u laboratoriji za proučavanje procesa mineralizacije kostiju i mogu se uzgajati (čuvati) izvan organizma.[13][14]
Ganoidne krljušti
uredi_(3149758934).jpg)

Ganoidne krljušti nalaze se u jesetri, vesola, garova, pramčanih perajarki, i sl. Izvedene su iz kosmoidnih krljušti i često imaju nazubljene ivice. Prekriveni su slojem tvrde cakline dentina na mjestu kosmina i slojem neorganske koštane soli zvane ganoin, umjesto vitrodentina.
Ganoin je karakteristična komponenta ganoidnih krljušti. To je staklasto, često višeslojno mineralizirano tkivo koje prekriva krljušti, kao i kranijalne kosti i zrakastih ribljih peraja u nekim neteleostnim ribama s perajima,[15] kao što su neke sjevernoameričke ribice Coelacanthi.[16][17] Sastoje se od štapićastih apatitnih kristalita.[18] Ganoin je drevno obilježje zrakastih peraja, a nalazi se naprimjer na krljuštims predaka aktinopterigijske Cheirolepis.[17] Iako se često smatra sinapomorfnimom karakter zrakoperskih riba, ganoin ili tkiva slična ganoinu također se nalaze na izumrlim Acanthodii.[17] Pretpostavlja se da je ganoin homologan sa zubnom caklinom kod kičmenjaka[15] ili se čak smatra tipom cakline.[18]
 Amblypterus striatus
|
Ganoidne krljušti izumrle karbonske ribe, Amblypterus striatus. (a) prikazuje vanjsku površinu četiri krljušti, a (b) unutrašnju površinu dvije krljušti. Svaka od ganoidnih krljušti romboidnog oblika roda Amblypterus ima na unutrašnjoj površini greben koji se na jednom kraju stvara u izbočeni klin koji se uklapa u zarez u sljedećoj skali, slično načinu na koji su pločice pričvršćene zajedno na krovu kuće. | 
|
Većina ganoidnih krljušti su romboidne (u obliku dijamanta) i povezane su spojevima klinova i utičnica. Obično su debele i spajaju se više kao ubodna pila, a ne preklapaju se kao druge krljušti.[19] Na ovaj način, ganoidne krljušti su gotovo neprobojne i odlična su zaštita od grabežljivaca.
-
![Aligatoroliki gar ima čvrst oklop od romboidnih ganoidnih krljušti.[19]](//upload.wikimedia.org/wikipedia/commons/thumb/b/bd/Alligator_gar_fish.jpg/395px-Alligator_gar_fish.jpg)
-

-
 Ganoidne ljuske na pramčanoj peraji su smanjene po veličini i podsjećaju na cikloidne krljušti.
Ganoidne ljuske na pramčanoj peraji su smanjene po veličini i podsjećaju na cikloidne krljušti.
![Aligatoroliki gar ima čvrst oklop od romboidnih ganoidnih krljušti.[19]](/wiki/Datoteka:Alligator_gar_fish.jpg)
.png)


Kod jesetri, krljušti su uveliko uvećane u oklopne ploče duž bočnih i stražnjih strana, dok je kod pramčanog peraja krljušt jako smanjena u debljini kako bi podsjećala na cikloidne krljušti.
-
 Naušnice napravljene od ganoidnih krljušti aligatorskog gara
Naušnice napravljene od ganoidnih krljušti aligatorskog gara -
 Fosil primitivne zrakoperke sa ganoidnim krljuštima
Fosil primitivne zrakoperke sa ganoidnim krljuštima -
 Ganoidne krljušti na fosiliziranom Lepidotes, prije oko. 130 miloona godina
Ganoidne krljušti na fosiliziranom Lepidotes, prije oko. 130 miloona godina


.jpg)
Starosjedioci Sjedinjenih Država i Karibljani koristili su čvrste ganoidne krljušti aligatorolikog gara za vrhove strela, prsne oklope i kao zaštitu za pokrivanje plugova. U današnje vrijeme od ovih vaga se pravi nakit.[20]
Leptoidne krljušti
urediLeptoidne (koštane krljušti) nalaze se na košljoribama višeg reda, teleosteama (što je izvedeniji kladus riba s perajima). Spoljašnji dio ovih krljušti je raširen sa koštanim grebenima, dok je unutrašnji dio isprepleten vlaknastim vezivnim tkivom. Leptoidne krljušti su tanje i prozirnije od drugih tipova i nemaju očvrsnute slojeve nalik na gleđ ili dentin. Za razliku od ganoidnih krljušti, dodatne ljuske se dodaju u koncentričnim slojevima, kako riba raste.[21]
Leptoidne krljušti se preklapaju u konfiguraciji od glave do repa, poput crijepa, što ih čini fleksibilnijim od kosmoidnih i ganoidnih. Ovaj raspored omogućava glatkiji protok vode preko tijela i smanjuje povlačenje.[22] Krljušti nekih vrsta pokazuju trake neujednačenog sezonskog rasta koje se nazivaju anuli (singular anulus). Ove trake se mogu koristiti za određivanje starosti ribe.
Leptoidne krljušti dolaze u dva oblika: cikloidne (glatke) i ktenoidne (češljaste).[23]
Cikloidne krljušti
urediCikloidne (kružne) krljušti imaju glatku teksturu i ujednačene su, sa glatkim vanjskim rubom ili obrubom. Najčešći su kod riba s mehkim perajama, kao što su losos i šaran.

|
.jpg)
|
Azijske arovane imaju velike cikloidne krljušti raspoređene na ribi u mozaiku podignutih rebara (lijevo). Same krljušti su prekrivene delikatnim mrežastim uzorkom (desno).[24][25]
| |


Cikloidne (kružne) ljuske obično se nalaze na šaranolikim ribama ili lososu. |







Ktenoidne krljušti
urediKtenoidne (nazubljene) krlušti su poput cikloidnih, osim što imaju male zupce ili spinule zvane kteni duž svojih vanjskih ili stražnjih rubova. Zbog ovih zuba krljušti imaju grubu teksturu. Obično se nalaze na ribama sa bodljikavom perajama, kao što su ribe smuđevi. Ove krljušti gotovo da ne sadrže kost, jer se sastoje od površinskog sloja koji sadrži hidroksiapatit i kalcij-karbonat i dubljeg sloja koji se sastoji uglavnom od kolagena. Caklina ostalih tipova ljuski je reducirana na površinske izbočine i ktenije.



|

|
Veličina zupca na ktenoidnim krljuštima može varirati u zavisnosti od položaja, kao što pokazuju ove kod Cetonurus crassiceps.
| |

_by_M._L._Nievera_(colored).png)
Ktenoidne (nazubljene) ljuske obično se nalaze na smuđolikim ribama.
|





Ktenoidne krljušti, slične drugim epidermnim strukturama, potiču od plakoidnih i karakteristična ćelijska diferencijacija ih čini isključivim iz drugih struktura koje nastaju iz integumenta.[27] Razvoj počinje blizu repnog perajq, duž bočne linije ribe.[28] Proces razvoja počinje akumulacijom fibroblasta između epidermisa i dermisa.[27] Kolagenske fibrile počinju da se organizuju u dermni sloj, što dovodi do inicijacije mineralizacije.[27] Prvo raste obim krljušti, a zatim debljina kada se slojevi koji se preklapaju zajedno mineraliziraju.[27]
Ktenoidne ljuske se dalje mogu podijeliti u tri tipa:
- Krenatne krljušti, gdje margina ima udubljenja i projekcije.
- Spinoidne krljušti, gdje krljušt nosi bodlje koje su kontinuirane sa samom krljušti.
- Prave ktenoidne krljušti, gdje su bodlje različite strukture.
Većina riba sa zračnim perajama ima ktenoidne krljušti. Neke vrste raža imaju ktenoidne krljušti na strani očiju i cikloidne na slijepoj strani, dok druge vrste imaju ktenoidne krljušti kod mužjaka i cikloidne kod ženki.
Refleksija
uredi


Mnoge košljoribe prekrivene su krljuštima koje se jako reflektiraju, koje funkcioniraju kao mala ogledala i daju izgled posrebrenog stakla. Refleksija srebrenjem je široko rasprostranjena ili dominantna kod riba otvorenog mora, posebno onih koje žive u gornjih 100 metara. Efekt transparentnosti može se postići posrebrenjem kako bi se tijelo životinje učinilo visoko reflektirajućim. Na srednjim dubinama na moru svjetlost dolazi odozgo, tako da vertikalno orijentirano ogledalo čini životinje poput riba nevidljivima sa strane.[29]
Morska sjekira je izuzetno spljoštena bočno (s jedne na drugu stranu), ostavljajući tijelo debljine samo milimetara, a tijelo je toliko srebreno da podsjeća na aluminijsku foliju. Ogledala se sastoje od mikroskopskih struktura sličnih onima koje se koriste za strukturno obojenje: hrpe od 5 do 10 kristala guanina razmaknute su oko ¼ talasne dužine da bi konstruktivno interferirale i postigle skoro 100 % refleksije. U dubokim vodama u kojima živi sjekira, samo plavo svjetlo s talasnom dužinom od 500 nanometara prodire i mora se reflektirati, tako da ogledala na udaljenosti od 125 nanometara pružaju dobru kamuflažu.[29]
Većina riba u gornjem dijelu oceana kamuflirana je posrebrenjem. Kod riba kao što je haringa, koja živi u plićoj vodi, ogledala moraju odražavati mješavinu talasnih dužina, a riba shodno tome ima kristalne hrpe s rasponom različitih razmaka. Daljnja komplikacija za ribe sa zaobljenim tijelima u poprečnom presjeku je to što ogledala ne bi bila efikasna ako bi se postavila ravno na kožu, ne bi reflektirala horizontalno. Ukupni efekt ogledala postiže se sa mnogo malih reflektora, svi su orijentisani okomito.[29]
Riblje krljušti sa ovim svojstvima koriste se u nekim kozmetičkim proizvodima, jer mogu dati svjetlucavi efekt šminki i ruževima.[30]
Plakoidne krljušti
uredi
Plakoidne (šiljaste, zubaste) krljušti se nalaze u hrskavičnih riba: ajkula, zrakaša. Nazivaju se i dermni zubići. Plakoidne krljušti su strukturno homologne sa pršljenom i zubima ("dentik" se prevodi kao "mali zub"), imaju središnju pulpnu šupljinu snabdjevenu krvnim sudovima, okruženu konusnim slojem dentina, a sve to leži na vrhu pravougaone bazne ploče koja se naslanja na dermis. Spoljni sloj se sastoji od vitrodentina, uglavnom neorganske supstance nalik caklini. Plakoidne krljušti ne mogu rasti u veličini, već se dodaje više ljuski kako se riba povećava.
Slične krljušti mogu se naći i ispod glave zubaste haringe. Količina pokrivenosti krljuštima je mnogo manja u zracima.
Sumnja se da romboidne krljušti sa svojstvima plakoidnih i ganoidnih ljuski postoje kod modernih predaka riba viličnjača: bezvilične Ostracoderma, a zatim sa vilicama Plakoderma.
Koža ajkule
uredi
Koža ajkule gotovo je u potpunosti prekrivena malim plakoidnim krljuštima. Potpomognute su bodljama, koje su grube kada se pomiču u smjeru unazad, ali kada se spljošte kretanjem vode naprijed, stvaraju sićušne vrtloge koji smanjuju hidrodinamički povlačenje i smanjuje turbulenciju, čineći plivanje efikasnijim i tišim u odnosu na košljoribe.[31] Takođe ima ulogu u sprečavanju obrastanja pokazujući lotosov efekt.[32]
Sve dentikule sastoje se od unutrašnje šupljine pulpe sa nervnim i arterijskim dovodom ukorijenjenim u dermis za opskrbu zubne sluzi.[33] Dentikule sadrže rebraste strukture koje strše iz površine krljušti; pod mikroskopom ovo rebro može izgledati kao udica ili grebeni koji izlaze iz krljušti. Ukupni oblik izbočine iz zuba ovisi o vrsti ajkule i općenito se može opisati s dva izgleda.[34] Prvi je krljušt u kojoj su grebeni postavljeni bočno niz ajkulu i paralelno sa tokom vode. Drugi oblik je glatka krljušt sa nečim što izgleda kao kukasti rebro koje se uvija iz površine i cilja prema stražnjoj strani ajkule.[34] Oba oblika rebraste forme pomažu u stvaranju turbulentnog graničnog sloja koji tjera laminarni tok dalje od kože morskog psa.[35]
Za razliku od košljoriba, ajkule imaju komplikovan dermni korzet napravljen od fleksibilnih kolagenskih slojevitih vlakana raspoređenih kao heliksna mreža koja okružuje njihovo tijelo. Korzet radi kao vanjski kostur, osiguravajući pričvršćivanje njihovih plivačkih mišića i na taj način štedi energiju.[36] Ovisno o položaju ovih plakoidnih krljišti na tijelu, one mogu biti fleksibilne i pasivno podignute, što im omogućava da mijenjaju svoj ugao napada. Također imaju rebra koja su poredana u smjeru toka, te rebraste trake smanjuju silu otpora koja djeluje na kožu ajkule tako što gura vrtlog dalje od površine kože, inhibirajući bilo kakav poprečni tok velike brzine.[37]
Morfologija krljušti
urediOpća anatomija krljušti varira, ali sve se mogu podijeliti na tri dijela: krunu, vrat i bazu. Savitljivost krljušti je povezana sa veličinom njene osnove. Krljušti veće fleksibilnosti imaju manju osnovu, pa su manje kruto vezane za stratum laxum. Na tjemenu brzoplivajućih ajkula nalazi se niz paralelnih rebara ili grebenova koji se protežu od prednje prema stražnjoj strani.[38]
Analizirajući tri komponente krljušti može se zaključiti da baza dentikula ne dolazi u kontakt ni sa jednim dijelom protoka tekućine.[39] Kruna i vrat zubaca, međutim, imaju ključnu ulogu i odgovorni su za stvaranje turbulentnih vrtloga i vrtloga koji se nalaze u blizini površine kože. Budući da zubići dolaze u toliko različitih oblika i veličina, može se očekivati da svi oblici neće proizvesti isti tip turbulentnog toka. Tokom nedavnog istraživačkog eksperimenta biomimetrijski uzorci zubaca ajkule sa mikrostrukturom poput polumjeseca i testirani u rezervoaru za vodu koristeći sto za vuču kao klizač. Eksperiment je pokazao da je površina sa zubima doživjela smanjenje otpora za 10% u odnosu na glatki uzorak. Razlog za ovo smanjenje otpora je taj što su turbulentni vrtlozi postali zarobljeni između zubaca, stvarajući barijeru poput jastuka protiv laminskog toka.[40] Ovaj isti tip eksperimenta izvela je druga istraživačka grupa koja je uvela više varijacija u svoj biometrijski uzorak. Druga grupa je došla do istog zaključka kao i prva, međutim, budući da je njihov eksperiment sadržavao više varijacija unutar uzoraka, uspjeli su postići visok stupanj eksperimentalne točnosti. U zaključku su naveli da su praktičniji oblici izdržljiviji od onih sa zamršenim linijama grebena. Praktični oblici su bili niskog profila i sadržavali su trapezoidne ili polukružne poprečne presjeke nalik na korita, i bili su manje efikasni, ali su ipak smanjili otpor za 6 ili 7%.[41]
Smanjenje povlačenja
uredi
Morski psi smanjuju otpor i ukupni trošak transporta (COT) kroz više različitih puteva. Otpor pritiska nastaje iz razlike u pritisku između prednje i zadnje strane ajkule zbog količine zapremine koja se gurne pored ajkule da bi se pokrenula naprijed.[42] Ovaj tip otpora je također direktno proporcionalan laminskom toku. Kada se laminski tok poveća oko ribe, otpor pritiska se također povećava.<r.[43] Otpor trenja rezultat je interakcije između tekućine i kože ajkule i može varirati ovisno o tome kako se granični sloj mijenja u odnosu na površinu ribe.[42]
Rebarca ometaju translaciju strujnih vrtloga u viskoznom podsloju. Mehanizam je složen i još nije u potpunosti shvaćen. U osnovi, inhibiraju formiranje vrtloga blizu površine jer se vrtlog ne može uklopiti u doline koje formiraju. Ovo gura vrtlog dalje od površine, u interakciji samo sa vrhovima rebara, ne izazivajući tok velike brzine u dolinama. Budući da ovaj tok velike brzine sada djeluje samo sa vrhom rebrca, koji je vrlo mala površina, prijenos momenta koji uzrokuje otpor je sada mnogo manji nego prije, čime se efektivno smanjuje otpor. Također, ovo smanjuje fluktuacije brzine poprečnog toka, što također pomaže u prijenosu momenta.[38]
Nedavna istraživanja pokazala su da postoji režim prije i nakon propadanja u graničnom sloju uz zid gdje se podsloj zgušnjava opadajućom brzinom, a zatim se naglo raspada u turbulentne vrtloge, prije nego što se konačno uruši. Ovaj sistem je potpuno samoregulirajući i posreduje u ciklusu rasta i propadanja; vrtlozi se akumuliraju tokom perioda rasta i naglo se likvidiraju u Strouhalove nizove ukosnih vrtloga koji se podižu sa zida. Vrtlozi koji se podižu su ono što gura granični sloj van i dalje od površine morskog psa, što rezultira smanjenjem ukupnog otpora koji doživljava riba.[44]
Gruba tekstura kože ajkule i raže nalik brusnom papiru, zajedno sa njenom žilavošću, dovela je do toga da se vrednuje kao izvor sirove ajkulske kože, zvane šagren. Jedna od mnogih historijskih primjena šagrena od ajkule bila je pravljenje rukohvata za mač. Gruba tekstura kože koristi se također u japanskoj kuhinji za pravljenje rendala pod nazivom oroshiki, pričvršćivanjem komada kože ajkule na drvene daske. Mala veličina krljušti vrlo fino nariba hranu.
Razvoj krljušti
urediKrljušti se obično pojavljuju kasno u razvoju ribe. U slučaju zebrica, potrebno je 30 dana nakon oplodnje prije nego što se različiti slojevi potrebni za formiranje krljušti diferenciraju i organiziraju. Za to je potrebno da dođe do konsolidacije mezenhima, zatim da se indukuje morfogeneze i konačno do procesa diferencijacije ili kasne metamorfoze.[45][46]
- Konsolidacija mezenhima: Konsolidacija ili strukturiranje mezenhima nastaje tokom razvoja dermisa. Ovaj proces ovisi o tome da li je riba hrskava ili košljoriba. Kod hrskavičnih riba strukturiranje nastaje formiranjem dva sloja. Prvi je površan i širok, a drugi tanak i kompaktan. Ova dva sloja su odvojena mezenhimske ćelijes. Koštane ribe stvaraju besćelijski supstrat organiziran okomito pomoću kolagenskih vlakana. Nakon toga, za obje ribe fibroblasti se izdužuju. Oni prodiru u kompaktni sloj mezenhima, koji se konsoliduje prije formiranja krljušti, kako bi pokrenuo dermnu ploču.[45][46][47]
- Indukcija morfogeneze: Morfogeneza je posljedica formiranja epidermne papile, koja nastaje spajanjem epiderme i dermisa kroz proces invaginacije. Morfogeneza počinje u vrijeme kada se fibroblasti presele u gornji dio kompaktnog mezenhima. Tokom ovog procesa, bazne ćelije epitela formiraju granični sloj, koji se nalazi u gornjem dijelu mezenhima. Nakon toga, ove ćelije će se diferencirati u području gdje će nastati primordij krljušti.[45][46][47]
- Diferencijacija ili kasna metamorfoza: Ovu diferencijaciju stvaraju dva različita oblika prema tipu krljušti koja se formira. Formiranje elazmoidnih krljušti (cikloidi i ktenoidi) nastaje stvaranjem prostora između matriksa epidermne papile. Ovaj prostor sadrži kolagenska vlakna. Oko ovog prostora diferenciraju se elazmoblasti i odgovorni su za stvaranje potrebnog materijala za formiranje krljušti. Nakon toga dolazi do matrikssne mineralizacije, omogućavajući krljuštima da steknu krutost koja ih identificira.[45][46][47]
Za razliku od elazmoidnih krljušti, ganoidne krljušti se sastoje od mineraliziranog i nemineraliziranog kolagena u različitim regijama. Formiranje ovih nastaje ulaskom površinskih ćelija mezenhima u matriks; potonji se sastoji od kolagenskih vlakana i nalazi se oko vaskularnih kapilara, stvarajući tako vaskularne šupljine. U ovoj fazi, elazmoblasti su zamijenjeni osteoblastima, čime se formira kost. Dijelovi matriksa krljušti koji nisu okoštani sastavljeni su od zbijenog kolagena koji mu omogućava da održi sjedinjenje s mezenhimom. Ovo je poznato kao Sharpeyeva vlakna.[45][46][47]
Jedan od gena koji reguliraju razvoj krljušt u ribama je zvučni jež (shh), koji je pomoću (shh) proteina uključen u organogenezu i u proces ćelijskih komunikacija, omogućavajući formiranje krljušti.[48][49] Apolipoprotein E (ApoE), koji omogućava transport i metabolizam triglicerida i holesterola, ima interakciju sa shh, jer ApoE obezbjeđuje holesterolni shh signalni put. Pokazalo se da je tokom procesa diferencijacije i interakcija ćelija nivo ApoE transkripcije visok, što je dovelo do zaključka da je ovaj protein važan za kasni razvoj krljušti.[48][49]
Modificirane krljušti
uredi



Različite grupe riba evoluirale su brojne modificirane krljušti za različite funkcije.
- Gotovo sve ribe imaju bočnu liniju, sistem mehanoreceptora koji detektuju kretanje vode. Kod košljoriba, krljušti duž bočne linije imaju centralne pore koje omogućavaju vodi da kontaktira senzorne ćelije.
- Bodlje leđnih peraja psećjih ajkula i himera, bodlje repa raža i "pilasti" zubi testerača i pilastih ajkula spojene su i modificirane plakoidne krljušti.
- Riba-hirurg ima oštricu nalik skalpelu, koja je modificirana krljušt, sa obje strane repnog pedunkula.[50]
- Neke haringe, inćuni i polukljun imaju listopadne ljuske, koje se lahko odbacuju i pomažu u bijegu od predatora.
- Strelice kod muških Percinae imaju red uvećanih kadukoznih krljušti između karličnog peraja i anusa.
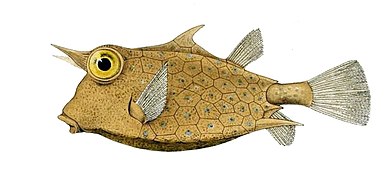
- Ribe-dikobrazi imaju krljušt modificiranu u velike vanjske kičme.
- Nasuprot tome, pufer-riba ima tanje, skrivenije bodlje od riba-dikobraza, koje postaju vidljive tek kada se riba napuhne.Za razliku od dikobraza, ove bodlje nisu modificirane krljušti, već se razvijaju pod kontrolom iste mreže gena koji proizvode perje i dlake kod drugih kičmenjaka.[51][52]
-
 Dikobraz-ribe imaju krljušti modificirane u kičmu.
Dikobraz-ribe imaju krljušti modificirane u kičmu. -
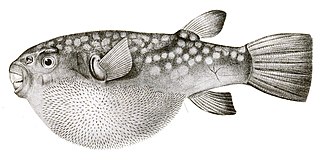 Pufer-riba ima bodlje koje nisu modificirane krljušti već su razvijene nezavisnom genskom mrežom.
Pufer-riba ima bodlje koje nisu modificirane krljušti već su razvijene nezavisnom genskom mrežom.


Ribe bez krljušti
uredi-
 Mandarinke nemaju krljušti i štite se slojem smrdljive i gorke sluzi.
Mandarinke nemaju krljušti i štite se slojem smrdljive i gorke sluzi.

Ribe bez krljušti obično razvijaju alternativu zaštiti koju krljušti mogu pružiti, kao što su tvrdi kožasti omotač ili koštane ploče.
- Bezviličnjače (lampetra i kolouste) imaju glatku kožu bez krljušti i bez kožnih kostiju.[53] Lampetre dobijaju određenu zaštitu od čvrste kože. Kolouste izlučuju velike količine sluzi ili sluz ako su ugrožene.[54] Mogu se vezati u čvor, stružući sluz dok idu i oslobađajući se od grabežljivca.[55]
- Većina jegulja je bez krljušti, iako su neke vrste prekrivene sitnim glatkim cikloidnim ljuskama.
- Većini somova nedostaju krljušti, iako nekoliko porodica ima pancire u obliku kožnih ploča ili neke tipove egzoskeleta.[56]
- Mandarinke nemaju krljušti i imaju sloj smrdljive i gorke sluzi koja blokira bolesti i vjerovatno obeshrabruje grabežljivce, što implicira da je njihova svijetla boja aposemijska.[57]
- Udičarke imaju labavu, tanku kožu često prekrivenu finim račvastim kožnim bodljama ili tuberkulima, ali nemaju pravilne krljušti. Oslanjaju se na kamuflažu kako bi izbjegli pažnju grabežljivaca, dok njihova labava koža otežava grabežljivcima da ih zgrabe.
Mnoge grupe košljoriba, uključujući iglice, morske konjiće, bokserske ribe, lovolovce, i nekoliko drugih porodica, razvile su vanjske koštane ploče, strukturno nalik plakoidnim krlčjuštima, kao zaštitni oklop od predatora.
- Morski konjići nemaju krljušti, ali imaju tanku kožu nategnutu preko koštanog pločastog oklopa raspoređenog u prstenove po dužini njihovog tijela.
- Kod bokserske ribe, ploče se spajaju i formiraju čvrstu školjku ili egzoskelet koji obuhvata cijelo tijelo. Ove koštane ploče nisu modificirane krljušti već koža koja je okoštala. Zbog ovog teškog oklopa ribice su ograničene na spore pokrete, ali malo drugih riba može pojesti odrasle jedinke.

| |
Jegulje izgledaju kao da su bez krljušti, ali neke vrste su prekrivene sitnim glatkim cikloidnim krljuštima.
| |
-
 Boserske ribe imaju ploče od okoštale kože spojene zajedno da formiraju čvrstu školjku.
Boserske ribe imaju ploče od okoštale kože spojene zajedno da formiraju čvrstu školjku. -
 Morski konjići imaju tanku kožu zategnutu preko koštanih ploča raspoređenih u prstenove.
Morski konjići imaju tanku kožu zategnutu preko koštanih ploča raspoređenih u prstenove.


-
Riba-kutija ima ploče od okoštale kože spojene zajedno da formiraju čvrstu školjku.
-
Morski konjići imaju tanku kožu zategnutu preko koštanih ploča raspoređenih u prstenove.
Neke ribe, poput hokija i sabljarke, rađaju se s krljuštima, ali ih odbacuju kako rastu.
Turpijaste ribe imaju grube krljušti koje se ne preklapaju sa malim šiljcima, zbog čega se tako nazivaju. Neke ribe izgledaju bez krljušti jer su im tako male.
Istaknuta krljušt pojavljuje se na tuni samo duž bočne linije i u korzeletu, zaštitnoj vrpci zadebljanih i uvećanih krljušti u predjelu ramena. Na većem dijelu tijela, tuna ima tako male ljuske da se na povremenim pregledom čini kao bez krljušti.[58]
-
 Neke turpijaste ribe izgledaju bez krljušti jer su im tako male.
Neke turpijaste ribe izgledaju bez krljušti jer su im tako male. -
 Slučajnom pregledu tuna izgleda uglavnom bez krljušti, ali nije.
Slučajnom pregledu tuna izgleda uglavnom bez krljušti, ali nije.


Levitikus
urediOdlomak u Levitikusu objavljuje da će "od svih što je u vodama... u morima i u rijekama" oni koji nemaju peraje krljušti "biti odvratni za vas" i mogu da ne se jedu.Greška kod citiranja: Nedostaje oznaka za zatvaranje </ref> nakon <ref> Definicija "krljušti" razlikuje se od definicija predstavljenih u biologiji, po tome što krljušti košer ribe moraju biti vidljive oku i mogu se lahko ukloniti sa kože bilo rukom ili nožem za ljuštenje.[59] Prema agenciji za sertifikaciju košera Pravoslavne unije, riba je košer ako se krljušt može ukloniti bez kidanja njene kože.[60] Tako su šaran i losos košer, dok ajkula, čija je krljušt mikroskopska, jesetra, čije se krljušti ne mogu lahko ukloniti a da se ne iseku iz tijela, nisu košer. Ostale nekošer ribe uključuju soma, jegulje, slatkovodni bakalar, zmijske skuše i napuhane ribe.[59]
Lepidofagija
uredi
Lepidofagija (stgrč. jedenje krljušti) je specijalizirano ponašanje u hranjenju riba koje uključuje jedenje krljušti drugih riba.[62] Lepidofagija je nezavisno evoluirala u najmanje pet slatkovodnih i sedam morskih porodica.[63]
Riblje krljušti mogu biti hranljive, sadrže dermni dio i sloj sluzi bogate proteinima, osim slojeva keratina i cakline. Bogat su izvor kalcij-fosfata.[63] Međutim, energija potrošena da se napravi udarac u odnosu na količinu krljušti koja se troši po udaru postavlja ograničenje na veličinu lepidofagne ribe, i obično su mnogo manji od svog plijena.[63] Ponašanje u ishrani obično se razvija zbog nedostatka hrane i ekstremnih uslova okoline. Jedenje krljušti i kože koja ih okružuje daje nutrijente bogate proteinima koji možda nisu dostupni nigdje drugdje u niši.[64]
Riblje vilice normalno imaju bilateralnu simetriju. Izuzetak se javlja kod ciklide Perissodus microlepis koja jede krljušti. Čeljusti ove ribe se javljaju u dva različita morfološka oblika. Jedan morf ima vilicu uvijenu ulijevo, što mu omogućava da lakše jede krljušti na desnom boku svoje žrtve. Drugi morf ima vilicu zakrivljenu udesno, što olakšava jedenje krljušti na lijevom boku žrtve. Relativna brojnost ova dva morfa u populacijama je regulisana frekvencijski zavisnom selekcijom.[61][65][66]
Također pogledajte
urediReference
uredi- ^ Scale Etymonline. Retrieved 28 April 2019.
- ^ Mongera, A.; Nüsslein-Volhard, C. (2013). "Scales of fish arise from mesoderm". Current Biology. 23 (9): R338–R339. doi:10.1016/j.cub.2013.02.056. PMID 23660349.
- ^ Sharpe, P. T. (2001). "Fish scale development: Hair today, teeth and scales yesterday?". Current Biology. 11 (18): R751–R752. doi:10.1016/S0960-9822(01)00438-9. PMID 11566120. S2CID 18868124.
- ^ Turner, S.; Tarling, D. H. (1982). "Thelodont and other agnathan distributions as tests of Lower Paleozoic continental reconstructions". Palaeogeography, Palaeoclimatology, Palaeoecology. 39 (3–4): 295–311. Bibcode:1982PPP....39..295T. doi:10.1016/0031-0182(82)90027-X.
- ^ Märss, T. (2006). "Exoskeletal ultrasculpture of early vertebrates". Journal of Vertebrate Paleontology. 26 (2): 235–252. doi:10.1671/0272-4634(2006)26[235:EUOEV]2.0.CO;2. S2CID 85993241.
- ^ a b Janvier, Philippe (1998). "Early vertebrates and their extant relatives". Early Vertebrates. Oxford University Press. str. 123–127. ISBN 978-0-19-854047-2.
- ^ a b Turner, S. (1991). "Monophyly and interrelationships of the Thelodonti". u M. M. Chang; Y. H. Liu; G. R. Zhang (ured.). Early Vertebrates and Related Problems of Evolutionary Biology. Science Press, Beijing. str. 87–119.
- ^ Märss, T. (1986). "Squamation of the thelodont agnathan Phlebolepis". Journal of Vertebrate Paleontology. 6 (1): 1–11. doi:10.1080/02724634.1986.10011593.
- ^ Botella, H.; J. I. Valenzuela-Rios; P. Carls (2006). "A New Early Devonian thelodont from Celtiberia (Spain), with a revision of Spanish thelodonts". Palaeontology. 49 (1): 141–154. doi:10.1111/j.1475-4983.2005.00534.x. S2CID 128939911.
- ^ Ferrón, Humberto G.; Botella, Héctor (2017). "Squamation and ecology of thelodonts". PLOS ONE. 12 (2): e0172781. Bibcode:2017PLoSO..1272781F. doi:10.1371/journal.pone.0172781. PMC 5328365. PMID 28241029.
- ^ MICHAEL ALLABY "cosmoid scale ." A Dictionary of Zoology . . Encyclopedia.com. 29 Oct. 2019 <https://www.encyclopedia.com>
- ^ Zylberberg, L., Meunier, F.J., Laurin, M. (2010). A microanatomical and histological study of the postcranial dermal skeleton in the Devonian sarcopterygian Eusthenopteron foordi, Acta Palaeontologica Polonica 55: 459–470.
- ^ Bergen, Dylan J. M.; Kague, Erika; Hammond, Chrissy L. (2019). "Zebrafish as an Emerging Model for Osteoporosis: A Primary Testing Platform for Screening New Osteo-Active Compounds". Frontiers in Endocrinology (jezik: engleski). 10: 6. doi:10.3389/fendo.2019.00006. ISSN 1664-2392. PMC 6361756. PMID 30761080.
- ^ de Vrieze, E.; van Kessel, M. A. H. J.; Peters, H. M.; Spanings, F. A. T.; Flik, G.; Metz, J. R. (1. 2. 2014). "Prednisolone induces osteoporosis-like phenotype in regenerating zebrafish scales". Osteoporosis International (jezik: engleski). 25 (2): 567–578. doi:10.1007/s00198-013-2441-3. ISSN 1433-2965. PMID 23903952. S2CID 21829206.
- ^ a b Zylberberg, L.; Sire, J. -Y.; Nanci, A. (1997). "Immunodetection of amelogenin-like proteins in the ganoine of experimentally regenerating scales of Calamoichthys calabaricus, a primitive actinopterygian fish". The Anatomical Record. 249 (1): 86–95. doi:10.1002/(SICI)1097-0185(199709)249:1<86::AID-AR11>3.0.CO;2-X. PMID 9294653.
- ^ Sire, Jean-Yves; Donoghue, Philip C. J.; Vickaryous, Matthews K. (2009). "Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates". Journal of Anatomy (jezik: engleski). 214 (4): 409–440. doi:10.1111/j.1469-7580.2009.01046.x. ISSN 0021-8782. PMC 2736117. PMID 19422423.
- ^ a b c Richter, M. (1995). "A microstructural study of the ganoine tissue of selected lower vertebrates". Zoological Journal of the Linnean Society. 114 (2): 173–212. doi:10.1006/zjls.1995.0023.
- ^ a b Bruet, B. J. F.; Song, J.; Boyce, M. C.; Ortiz, C. (2008). "Materials design principles of ancient fish armour". Nature Materials. 7 (9): 748–756. Bibcode:2008NatMa...7..748B. doi:10.1038/nmat2231. PMID 18660814.
- ^ a b Sherman, Vincent R.; Yaraghi, Nicholas A.; Kisailus, David; Meyers, Marc A. (1. 12. 2016). "Microstructural and geometric influences in the protective scales of Atractosteus spatula". Journal of the Royal Society Interface (jezik: engleski). 13 (125): 20160595. doi:10.1098/rsif.2016.0595. ISSN 1742-5689. PMC 5221522. PMID 27974575.
- ^ "Missouri Alligator Gar Management and Restoration Plan" (PDF). Missouri Department of Conservation Fisheries Division. 22. 1. 2013. Arhivirano s originala (PDF), 6. 5. 2016. Pristupljeno 12. 4. 2019.
- ^ Lagler, K. F., J. E. Bardach, and R. R. Miller (1962) Ichthyology. New York: John Wiley & Sons.
- ^ Ballard, Bonnie; Cheek, Ryan (2. 7. 2016). Exotic Animal Medicine for the Veterinary Technician. John Wiley & Sons. ISBN 978-1-118-92421-1.
- ^ McGrouther, Mark (2 Dec 2019). "Cycloid and Ctenoid Scales". The Australian Museum. Pristupljeno 29. 12. 2021.
- ^ Pouyaud, L.; Sudarto, Guy G. Teugels (2003). "The different colour varieties of the Asian arowana Scleropages formosus (Osteoglossidae) are distinct species: morphologic and genetic evidences". Cybium. 27 (4): 287–305.
- ^ Ismail, M. (1989). Systematics, Zoogeography, and Conservation of the Freshwater Fishes of Peninsular Malaysia (Doctoral Dissertation izd.). Colorado State University.
- ^ E.J. Brill (1953). The Fishes of the Indo-Australian Archipelago. E.J. Brill. str. 306–307.
- ^ a b c d Kawasaki, Kenta C., "A Genetic Analysis of Cichlid Scale Morphology" (2016). Masters Theses May 2014 - current. 425. http://scholarworks.umass.edu/masters_theses_2/425
- ^ Helfman, Gene (2009). The Diversity of Fishes Biology, Evolution, and Ecology. Wiley-Blackwell.
- ^ a b c Herring, Peter. The Biology of the Deep Ocean. Oxford: Oxford University Press. str. 193–195. ISBN 9780198549567.
- ^ "There Are Probably Fish Scales In Your Lipstick". HuffPost India (jezik: engleski). 23. 4. 2015. Pristupljeno 6. 5. 2019.
- ^ Martin, R. Aidan. "Skin of the Teeth". Pristupljeno 28. 8. 2007.
- ^ Fürstner, Reiner; Barthlott, Wilhelm; Neinhuis, Christoph; Walzel, Peter (1. 2. 2005). "Wetting and Self-Cleaning Properties of Artificial Superhydrophobic Surfaces". Langmuir. 21 (3): 956–961. doi:10.1021/la0401011. ISSN 0743-7463. PMID 15667174.
- ^ Lauder, George V.; Wainwright, Dylan K.; Domel, August G.; Weaver, James C.; Wen, Li; Bertoldi, Katia (2016). "Structure, biomimetics, and fluid dynamics of fish skin surfaces". Physical Review Fluids. 1 (6): 060502. Bibcode:2016PhRvF...1f0502L. doi:10.1103/PhysRevFluids.1.060502. S2CID 18118663.
- ^ a b Feld, Katrine; Kolborg, Anne Noer; Nyborg, Camilla Marie; Salewski, Mirko; Steffensen, John Fleng; Berg-Sørensen, Kirstine (24. 5. 2019). "Dermal Denticles of Three Slowly Swimming Shark Species: Microscopy and Flow Visualization". Biomimetics. 4 (2): 38. doi:10.3390/biomimetics4020038. ISSN 2313-7673. PMC 6631580. PMID 31137624.
- ^ Fletcher, Thomas; Altringham, John; Peakall, Jeffrey; Wignall, Paul; Dorrell, Robert (7. 8. 2014). "Hydrodynamics of fossil fishes". Proceedings of the Royal Society B: Biological Sciences. 281 (1788): 20140703. doi:10.1098/rspb.2014.0703. ISSN 0962-8452. PMC 4083790. PMID 24943377.
- ^ Martin, R. Aidan. "The Importance of Being Cartilaginous". ReefQuest Centre for Shark Research. Pristupljeno 29. 8. 2009.
- ^ Hage, W.; Bruse, M.; Bechert, D. W. (1. 5. 2000). "Experiments with three-dimensional riblets as an idealized model of shark skin". Experiments in Fluids (jezik: engleski). 28 (5): 403–412. Bibcode:2000ExFl...28..403B. doi:10.1007/s003480050400. ISSN 1432-1114. S2CID 122574419.
- ^ a b Motta, Philip; Habegger, Maria Laura; Lang, Amy; Hueter, Robert; Davis, Jessica (1. 10. 2012). "Scale morphology and flexibility in the shortfin mako Isurus oxyrinchus and the blacktip shark Carcharhinus limbatus". Journal of Morphology (jezik: engleski). 273 (10): 1096–1110. doi:10.1002/jmor.20047. ISSN 1097-4687. PMID 22730019. S2CID 23881820.
- ^ Dou, Zhaoliang; Wang, Jiadao; Chen, Darong (1. 12. 2012). "Bionic Research on Fish Scales for Drag Reduction". Journal of Bionic Engineering. 9 (4): 457–464. doi:10.1016/S1672-6529(11)60140-6. ISSN 1672-6529. S2CID 137143652.
- ^ "Experimental investigations on drag-reduction characteristics of bionic surface with water-trapping microstructures of fish scales" (PDF).
- ^ Palmer, Colin; Young, Mark T. (14. 1. 2015). "Surface drag reduction and flow separation control in pelagic vertebrates, with implications for interpreting scale morphologies in fossil taxa". Royal Society Open Science. 2 (1): 140163. Bibcode:2015RSOS....240163P. doi:10.1098/rsos.140163. ISSN 2054-5703. PMC 4448786. PMID 26064576.
- ^ a b Lauder, George V.; Wainwright, Dylan K.; Domel, August G.; Weaver, James C.; Wen, Li; Bertoldi, Katia (18. 10. 2016). "Structure, biomimetics, and fluid dynamics of fish skin surfaces". Physical Review Fluids. 1 (6): 060502. Bibcode:2016PhRvF...1f0502L. doi:10.1103/PhysRevFluids.1.060502.
- ^ Muthuramalingam, Muthukumar; Villemin, Leo S.; Bruecker, Christoph (29. 4. 2019). "Streak formation in flow over Biomimetic Fish Scale Arrays". The Journal of Experimental Biology (jezik: engleski). 222 (Pt 16): jeb205963. arXiv:1904.12752. Bibcode:2019arXiv190412752M. doi:10.1242/jeb.205963. PMID 31375542. S2CID 139103148.
- ^ Bandyopadhyay, Promode R.; Hellum, Aren M. (23. 10. 2014). "Modeling how shark and dolphin skin patterns control transitional wall-turbulence vorticity patterns using spatiotemporal phase reset mechanisms". Scientific Reports (jezik: engleski). 4: 6650. Bibcode:2014NatSR...4E6650B. doi:10.1038/srep06650. ISSN 2045-2322. PMC 4206846. PMID 25338940.
- ^ a b c d e Sire, J.Y.; Huysseune, A.N.N. (2003). "Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach". Biological Reviews. 78 (2): 219–249. doi:10.1017/S1464793102006073. PMID 12803422. S2CID 19556201.
- ^ a b c d e Le Guellec, D.; Morvan-Dubois, G.; Sire, J.Y. (2004). "Skin development in bony fish with particular emphasis on collagen deposition in the dermis of the zebrafish (Danio rerio)". International Journal of Developmental Biology. 48 (2–3): 217–231. doi:10.1387/ijdb.15272388. PMID 15272388.
- ^ a b c d Sire, J.Y. (2001). "Teeth outside the mouth in teleost fishes: how to benefit from a developmental accident". Evolution & Development. 3 (2): 104–108. doi:10.1046/j.1525-142x.2001.003002104.x. PMID 11341672. S2CID 13353402.
- ^ a b Sire, J.Y.; Akimenko, M.A. (2003). "Scale development in fish: a review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio)". International Journal of Developmental Biology. 48 (2–3): 233–247. doi:10.1387/ijdb.15272389. PMID 15272389.
- ^ a b Monnot, M.J.; Babin, P.J.; Poleo, G.; Andre, M.; Laforest, L.; Ballagny, C.; Akimenko, M.A. (1999). "Epidermal expression of apolipoprotein E gene during fin and scale development and fin regeneration in zebrafish". Developmental Dynamics. 214 (3): 207–215. doi:10.1002/(SICI)1097-0177(199903)214:3<207::AID-AJA4>3.0.CO;2-5. PMID 10090147.
- ^ Sorenson, L.; Santini, F.; Carnevale, G.; Alfaro, M.E. (2013). "A multi-locus timetree of surgeonfishes (Acanthuridae, Percomorpha), with revised family taxonomy". Molecular Phylogenetics & Evolution. 68 (1): 150–160. doi:10.1016/j.ympev.2013.03.014. PMID 23542000.
- ^ How the pufferfish got its wacky spines Phys.org, 25 July 2019.
- ^ Shono, T.; Thiery, A.P.; Cooper, R.L.; Kurokawa, D.; Britz, R.; Okabe, M.; Fraser, G.J. (2019). "Evolution and Developmental Diversity of Skin Spines in Pufferfishes". iScience. 19: 1248–1259. Bibcode:2019iSci...19.1248S. doi:10.1016/j.isci.2019.06.003. PMC 6831732. PMID 31353167.
- ^ Coolidge E, Hedrick MS and Milsom WK (2011) "Ventilatory Systems". In: McKenzie DJ, Farrell AP and Brauner CJ (Eds) Fish Physiology: Primitive Fishes, Elsevier, Page 182–213. ISBN 9780080549521
- ^ {{cite news | last=Rothschild | first=Anna | title=Hagfish slime: The clothing of the future? [[[[| url= https://www.bbc.co.uk/news/magazine-21954779 | work=BBC News | date= 1. 4. 2013 | access-date=2. 4. 2013}}
- ^ Yong, Ed (23. 1. 2019). "No One Is Prepared for Hagfish Slime". The Atlantic (jezik: engleski). Pristupljeno 26. 1. 2019.
- ^ Friel, J P; Lundberg, J G (1996). "Micromyzon akamai, gen. et sp. nov., a small and eyeless banjo catfish (Siluriformes: Aspredinidae) from the river channels of the lower Amazon basin". Copeia. 1996 (3): 641–648. doi:10.2307/1447528. JSTOR 1447528.
- ^ Sadovy, Y.; Randall, J. E.; Rasotto, Maria B. (maj 2005). "Skin structure in six dragonet species (Gobiesociformes; Callionymidae): Interspecific differences in glandular cell types and mucus secretion". Journal of Fish Biology. 66 (5): 1411–1418. doi:10.1111/j.0022-1112.2005.00692.x.
- ^ Do tunas have scales? Northeast Fisheries Science Center, NOAA Fisheries. Accessed 4 August 2019.
- ^ a b Greška kod citiranja: Nevaljana oznaka
<ref>; nije naveden tekst za reference s imenomchabad.org - ^ Verifying Kosher Fish OU Kosher Certification. Retrieved 9 August 2019.
- ^ a b Lee, H. J.; Kusche, H.; Meyer, A. (2012). "Handed Foraging Behavior in Scale-Eating Cichlid Fish: Its Potential Role in Shaping Morphological Asymmetry". PLOS ONE. 7 (9): e44670. Bibcode:2012PLoSO...744670L. doi:10.1371/journal.pone.0044670. PMC 3435272. PMID 22970282.
- ^ Froese, R. and D. Pauly. Editors. "Glossary: Lepidophagy". FishBase. Pristupljeno 12. 4. 2007.
- ^ a b c Janovetz, Jeff (2005). "Functional morphology of feeding in the scale-eating specialist Catoprion mento" (PDF). The Journal of Experimental Biology. 208 (Pt 24): 4757–4768. doi:10.1242/jeb.01938. PMID 16326957. S2CID 15566769.
- ^ Martin, C.; P.C. Wainwright (2011). "Trophic novelty is linked to exceptional rates of morphological diversification in two adaptive radiations of Cyprinodon pupfish". Evolution. 65 (8): 2197–2212. doi:10.1111/j.1558-5646.2011.01294.x. PMID 21790569. S2CID 23695342.
- ^ Hori, M. (1993). "Frequency-dependent natural selection in the handedness of scale-eating cichlid fish". Science. 260 (5105): 216–219. Bibcode:1993Sci...260..216H. doi:10.1126/science.260.5105.216. PMID 17807183. S2CID 33113282.
- ^ Stewart, T. A.; Albertson, R. C. (2010). "Evolution of a unique predatory feeding apparatus: functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids". BMC Biology. 8 (1): 8. doi:10.1186/1741-7007-8-8. PMC 2828976. PMID 20102595.
Dopunska literatura
uredi- Helfman, G.S., B.B. Collette and D.E. Facey (1997). The Diversity of Fishes. Blackwell Science. str. 33–36. ISBN 978-0-86542-256-8.CS1 održavanje: više imena: authors list (link)
- Schultze, H.P. (2016). "Scales, enamel, cosmine, ganoine, and early osteichthyans". Comptes Rendus Palevol. 15 (1–2): 83–102. doi:10.1016/j.crpv.2015.04.001.