Umrežavanje DNK
U genetici, umrežavanje DNK označava pojave kada različiti egzogeni ili endogeni agensi reaguju sa dva nukleotida DNK, formirajući kovalentnu vezu između njih. Ova poprečna veza može se desiti unutar istog lanca (unutarlančana) ili između suprotnih lanaca dvolančane DNK (međulančana). Ovi adukti ometaju ćelijski metabolizam, kao što su replikacija DNK i transkripcija, pokrećući ćelijsku smrt. Ove poprečne veze se, međutim, mogu popraviti putem ekscizije ili rekombinacijskih puteva.

Umrežavanje DNK također ima korisne zasluge u hemoterapijama i ciljanju kancerogenih ćelija za apoptoze,[1] kao i u razumijevanju interakcije proteina sa DNK.
Agensi umrežavanja
urediMnogi karakterizirani agensi za umrežavanje imaju dvije neovisno reaktivne grupe unutar iste molekule, od kojih se svaka može vezati za nukleotidni ostatak DNK. Ovi agensi su odvojeni na osnovu njihovog izvora i označeni kao egzogeni ili endogeni. Egzogeni agensi za umrežavanje su hemikalije i spojevi, prirodni i sintetski, koji proizlaze iz izloženosti okolišu kao što su lijekovi i dim cigareta ili izduvni gasovi automobila. Endogeni agensi za umrežavanje su spojevi i metaboliti koji se unose iz ćelijskih ili biohemijskih puteva unutar ćelije ili organizma.
Egzogeni agensi
uredi- Dušični iperit su egzogeni alkilirajući agensi koji reaguju sa N7 položajem guanina. Ovi spojevi imaju strukturu jezgra bis-(2-etilhloro)amina, sa promenljivom R-grupom, sa dvije reaktivne funkcionalne grupe koje služe za alkilaciju nukleobaza i formiraju umreženu leziju. Ovi agensi najpoželjnije formiraju 1,3 5'-d(GNC) međulančanu poprečnu vezu. Uvođenje ovog agensa blago savija DNK dupleks, kako bi se prilagodio prisutnosti agensa unutar heliksa.[2] Ovi agensi se često uvode kao farmaceutski proizvodi i koriste se u citotoksičnoj hemoterapiji.[3]
- Cisplatin (cis-diamindikloroplatina(II)) i njegovi derivati uglavnom deluju na susedne gvanine na njihovim N7 pozicijama. Planarno jedinjenje se povezuje s nukleobazama putem pomicanja vode jedne ili obje svoje kloridne grupe, omogućavajući cisplatinu da formira monoadukte za DNK ili RNK, unakrsne veze unutar lanaca DNK, umreženost međulančanih DNK i umrežavanje DNK-protein.[4] Kada cisplatin generira umrežavanje DNK, on češće formira 1,2-intralančane umrežene veze (5'-GG), ali također formira i 1,3-intralančane umrežene veze (5-GNG) u nižim procentima.[5][6] Kada cisplatin formira međulančano umrežavanje (5'-GC), dolazi do ozbiljnog izobličenja spirale DNK zbog skraćene udaljenosti između guanina na suprotnim nitima i citozina koji je izbačen iz heliksa kao posljedica interakcije GG.[7] Slično dušičnim iperitima, cisplatin se često koristi u liječenju hemoterapijom – posebno za rak testisa i jajnika.[8]
- Hloro-etil nitrozo-urea (CENU), posebno karmustin (BCNU), su agensi za umrežavanje koji se široko koriste u hemoterapiji, posebno za tumore mozga. Ovi agensi razlikuju se od drugih umreživača jer alkiliraju O6 guanin da bi formirali O6-etanoguanin. Ovaj intermedijarni spoj zatim dovodi do umrežavanja između baznog para GC. Ovi agensi za umrežavanje dovode samo do malih izobličenja spirale DNK zbog manje veličine molekula.
- Psoraleni su prirodni spojevi (furokumarini) u biljkama. Ovi spojevi interkaliraju u DNK na mestima 5'-AT sekvence i formiraju timidinske adukte kada se aktiviraju u prisustvu ultraljubičastih-A (UV-A) zraka.[9] Ovi kovalentni adukti nastaju povezivanjem 3, 4 (piron) ili 4', 5’ (furan) ruba psoralena sa 5, 6 dvostrukom vezom timina. Psoraleni mogu formirati dva tipa monoadukta i jedan dijadukt (međulančano umrežavanje) sa timinom.[10] Ovi adukti rezultiraju lokalnim distorzijama DNK na mjestu interkalacije. Psoraleni se koriste u liječenju kožnih bolesti, kao što su psorijaza i vitiligo.
- Mitomicin C (MMC) je iz klase antibiotika koji se široko koriste u hemoterapiji, često kod karcinoma povezanih sa gastrointestinalnim traktom. Mitomicin C može djelovati samo kao umreživač kada je DNK nukleotid imao redukciju na svom kinonskom prstenu. Kada su dva dG-a preuređena i metilirana na ovaj način, 5'-GC međulančano umrežavanje može se formirati sa egzo aminima svake nukleobaze. Mitomicin također ima sposobnost da formira monoadukte i međulančane poprečne veze sa DNK. Međulančana umrežavanja mitomicina C formiraju se u manjem žlijebu DNK, izazivajući umjereno širenje ili istezanje do spirale DNK kako bi se prilagodio prisustvu molekula unutar dva lanca.
Endogeni agensi
uredi- Dušična kiselina se formira kao nusprodukt u želucu iz prehrambenih izvora nitrita i može dovesti do umreženih lezija u DNK, konverzijom amino grupa u DNK u karbonile. Ovaj tip lezije najčešće se javlja između dva gvanozina, pri čemu jedan od četiri deaminirana gvanozina dovodi do međulančanog umrežavanja.[11] On indukuje formiranje međulančanih DNK ukrštenih veza na aminogrupi egzocikličkog N2 guanina na 5'-CG sekvencama. Ova lezija blago iskrivljuje dvostruku spiralu.
- Bifunkcionalni aldehidi su reaktivne hemikalije koje se formiraju endogeno putem peroksidacija lipida i biosinteze prostoglandina.[12] Oni stvaraju eteno adukte formirane od aldehida koji se podvrgavaju preuređenju kako bi formirali umrežavanje na suprotnim lancima DNK. Malondialdehid je prototipski primjer koji može umrežiti DNK preko dvije egzociklične guaninske amino grupe.[13] Otali aldehidi, kao što su formaldehid i acetilaldehidi, mogu uvesti međulančano umrežavanje i često djeluju kao egzogeni agensi jer se nalaze u mnogim prerađenim namirnicama. Često pronađeni u pesticidima, duhanskom dimu i automobilskim izduvnim plinovima, α,β nezasićeni aldehidi, kao što su akrolein i krotonaldehid, su dodatni egzogeni agensi koji mogu izazvati umrežavanje DNK. Za razliku od drugih agenasa za umrežavanje, umrežavanje izazvano aldehidom je suštinski reverzibilan proces. NMR struktura ovih tipova agenasa kao međulančanih poprečnih veza pokazuje da 5'-GC adukt rezultira manjim izobličenjem DNK, međutim 5'-CG adukt destabilizira spiralu i izaziva savijanje i uvijanje u DNK.[14]
- Umrežene lezije DNK mogu se formirati i u uslovima oksidativnog stresa, u kojem slobodni radikali kisika stvaraju reaktivne intermedijere u DNK, a ove lezije su uključene u starenje i rak. Tandemske lezije DNK formiraju se na značajnoj frekvenciji ionizirajućim zračenjem i reakcijama H2O2 kataliziranim metalom. U anoksijskim uslovima, dominantna lezija dvostruke baze je tip u kojoj je C8 guanin vezan za 5-metilnu grupu susjednog 3'-timina (G[8,5-Me]T), formirajući unutarlančane lezije.[15][16]
Zbirna tabela agenasa za umrežavanje
uredi| Umrežavajući agens | Alkilirajući agens | Struktura umreživača | Preferencijalna ciljna sekvenca |
| Dušični senf |   |
Unujtarlančano; 5'-GC | |
| Cisplatin |  |
 |
Unutarlančano; 5'-GC
Međulančano; 5'-GNG |
| Karmustin (BNCU) |  |
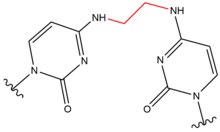 |
Međulančano; bazni par GC |
| Mitomicin C |  |
 |
Međulančano; 5'-GC |
| Psoralen |   |
Međulančano; 5'-TA | |
| Malondialdehid |  |
 |
Međulančano; 5'-GC, 5'-CG |
| Oksidativni stres (i zračrnje) |  |
Unutarlančano; d(GpT) | |
| Dušična kiselina |  |
 |
Međulančano; 5'-CG |
Popravak umrežene DNK
urediUmrežena DNK se popravlja u ćelijama kombinacijom enzima i drugih faktora iz puta nukleotidnog ekscizijskog popravka (NER), homolognom rekombinacijom i puta baznoekscijskog popravka (BER). Za popravak međulančanih umreženih veza kod eukariota, endonukleaza od 3' preklopa iz NER-a, XPF-ERCC1, regrutuje se u umreženu DNK, gdje pomaže u 'otkačivanju' DNK, cijepanjem 3' lanca na mjestu umrežavanja. 5' lanac se zatim cijepa, bilo pomoću XPF-ERCC1 ili druge endonukleaze, formirajući dvolančani prekid (DSB), koji se zatim može popraviti pomoću puta homologne rekombinacije.[17]
Umređena DNK općenito uzrokuju gubitak informacija o preklapanju sekvenci iz dva lanca DNK. Stoga, tačna popravka oštećenja ovisi o preuzimanju izgubljene informacije iz neoštećenog homolognog hromosoma u istoj ćeliji. Preuzimanje se može dogoditi uparivanjem sa sestrinskom hromatidom proizvedenom tokom prethodnog kruga replikacije. U diploidnoj ćeliji umrežavanje se takođe može desiti uparivanjem sa nesestrinskim homolognim hromosomom, što se dešava posebno tokom mejoze. Kada dođe do uparivanja, umrežavanje se može ukloniti i ispravne informacije uvesti u oštećeni hromosom, homolognom rekombinacijom.
Raskid veze između šećera dezoksiriboze u šećerno-fosfatnoj kičmi DNK i povezane nukleobaza ostavlja abazno mjesto u dvolančanoj DNK. Ova abazna mjesta se često stvaraju kao međuproizvodi, a zatim se obnavljaju u baznoj ekscizijskoj popravci. Međutim, ako se ovim mjestima dopusti da opstanu, mogu inhibirati replikaciju i transkripciju DNK.[18] Abazna mjesta mogu reagovati sa aminskim grupama na proteinima da bi formirali umrežavanje DNK-protein ili sa egzocikličkim aminima drugih nukleobaza da bi formirale međulančane umrežene veze. Kako bi spriječili međulančane ili umrežavanje DNK-protein, enzimi iz BER puta čvrsto vezuju abazno mjesto i izdvajaju ga iz obližnjih reaktivnih grupa, kao što je pokazano u ljudskoj alkiladenin DNK glikozilazi (AAG) i E. coli 3-metiladenin DNK glikozilazi II (AlkA).[19] In vitro dokazi pokazali su da međulančano umrežavanje inducirano nebaznim mjestom (DOB-ICL) predstavljaju lezije koje blokiraju replikaciju i vrlo pogrešno kodiraju. U poređenju sa nekoliko drugih ispitanih TLS pol-a, pol η će vjerovatno doprinijeti TLS-posredovanoj popravci DOB-ICL in vivo.[20] Korištenjem O6-2'-deoksigvanozin-butilen-O6-2'-deoksiguanozin (O6-dG-C4-O6-dG) lezija DNK koja je hemijski stabilna struktura, istraživana je aktivnost premošćavanja nekoliko DNK polimeraza i rezultati su pokazali da pol η ima najveću aktivnost premošćavanja; međutim, 70% bajpas proizvoda je bilo mutageno i sadrži zamjene ili delecije. Povećanje veličine nepovezanih popravnih intermedijera podiže učestalost delecijskih mutacija.[21]
Tretiranje E. coli sa psoralenom-plus-UV svjetlom (PUVA) stvara međulančano umredžavanje DNK. Cole et al.[22] i Sinden & Cole[23] predstavili su dokaz da homolognorekombinacijski proces popravke koji zahtijeva proizvode gena uvrA, uvrB i recA može ukloniti ova umrežavanja u E. coli. Čini se da je ovaj proces prilično efikasan. Iako su jedno ili dva nepopravljena umrećavanja dovoljna da inaktiviraju ćeliju, bakterijska ćelija divljeg tipa može se popraviti i stoga oporaviti od 53 do 71 psoralenskih umrežavanja. Eukariotske ćelije kvasca su također inaktivirane jednom preostalom umreženom vezom, ali ćelije kvasca divljeg tipa mogu se oporaviti od 120 do 200 umrežavanja.[24]
Aplikacije
urediUmrežavanje DNK i proteina
urediBiohemijski metodi interakcije
urediUmrežavanje DNK-protein može biti uzrokovano raznim hemijskim i fizičkim agensima, uključujući prijelazne metale, ionizirajuće zračenje i endogene aldehide, pored hemoterapeutskih agenasa.[25] eDNK umrežavanju, DNK-protein umrežavanju su lezije u ćelijama koje se često oštećuju UV zračenjem. Utjecaj UV zraka može dovesti do reaktivnih interakcija i uzrokovati umrežavanje DNK i proteina koji su u kontaktu s njom. Ove umrežavajuće veze su veoma glomazne i složene lezije. One se prvenstveno javljaju u područjima hromosoma koji prolaze kroz replikaciju DNK i ometaju ćelijske procese.
Napredak u metodima identifikacije strukture je napredovao, a dodatak sposobnosti mjerenja interakcija između DNK i proteina je zahtjev za potpuno razumijevanje biohemijskih procesa. Struktura kompleksa DNK-protein može se mapirati pomoću fotoumrežavanja, što je fotoindukovano formiranje kovalentne veze između dva makromolekula ili između dva različita dijela jedne makromolekule. Metodologija uključuje kovalentno povezivanje DNK-vezujućeg motiva DNK-vezujućeg proteina specifičnog za ciljnu sekvencu sa agensom za umrežavanje koji se može fotoaktivirati i koji je sposoban da reaguje sa nukleotidima DNK kada je izložen UV zračenju. Ovaj metoda daje informacije o interakciji između DNK i proteina u unakrsnoj vezi umrežavanja.[26]
Klinički tretmani
urediPutevi popravka DNK mogu dovesti do stvaranja tumorskih ćelija. Kancerski tretmani su dizajnirani pomoću agenasa za umrežavanje DNK za interakciju sa dušičnim bazama DNK kako bi se blokirala replikacija DNK. Ovi agensi za umrežavanje imaju sposobnost da djeluju kao terapije sa jednim agensom ciljajući i uništavajući specifične nukleotide u ćelijama raka. Rezultat je zaustavljanje ciklusa i rasta ćelija raka; budući da inhibira specifične puteve popravka DNK, ovaj pristup ima potencijalnu prednost jer ima manje nuspojava.[27]
Kod ljudi, vodeći uzrok smrti od raka u svijetu je rak pluća, uključujući karcinom nemalih a plućnih ćelija (NSCLC) koji čini 85% svih slučajeva raka pluća u Sjedinjenim Državama.[28] Pacijenti sa NSCLC liječe se često terapeutskim spojevima platine (npr. cisplatin, karboplatin ili oksaliplatin) koji uzrokuju međulančano umrežavanje DNK. Među osobama sa NSLC-om, niska ekspresija gena raka dojke 1 (BRCA1) u primarnom tumoru korelirala je s poboljšanim preživljavanjem nakon hemoterapije koja sadrži platinu.[29][30] Ova korelacija implicira da nizak BRCA1 u kanceru, i posljedično nizak nivo popravka DNK, uzrokuje ranjivost raka na liječenje agensima za umrežavanje DNK. Visok BRCA1 može zaštititi ćelije raka djelujući u homolognorekombinacijskom putu popravka koji uklanja oštećenja u DNK unesena lijekovima platine. Nivo ekspresije BRCA1 potencijalno je važan alat za prilagođavanje hemoterapije u liječenju raka pluća.[29][30]
Klinički hemoterapeutici mogu inducirati enzimsko i neenzimsko umrežavanje DNK-protein. Primjer ove indukcije su derivati platine, kao što su cisplatin i oksaliplatin. Oni stvaraju neenzimsko umrežavanje DNK-protein kroz nespecifično umrežavanje proteina koji stupaju u interakciju s hromatinom za DNK. Umrežavanje je moguće i kod drugih terapeutskih agenasa, bilo stabilizacijom intermedijara reakcije kovalentne DNK-protein ili stvaranjem pseudosupstrata, koji hvata enzim na DNK. Derivati kamptotecina, kao što su irinotekan i topotekan, ciljaju i hvataju specifičnu DNK topoizomerazu 1 (TOP1), interkalacijom unutar interfejsa enzim-DNK. Budući da toksičnost ovih lijekova ovisi o hvatanju TOP1, ćelijska osjetljivost na ove spojeve direktno zavisi od nivoa ekspresije TOP1. Kao rezultat toga, funkcija ovih lijekova je da služe kao otrovi enzima, a ne kao inhibitori. Ovo se može primijeniti za liječenje tumorskih ćelija, korištenjem TOP 2 otrova enzima.[31]
Reference
uredi- ^ Deans, AJ; West, SC (24. 6. 2011). "DNA interstrand crosslink repair and cancer". Nature Reviews. Cancer. 11 (7): 467–80. doi:10.1038/nrc3088. PMC 3560328. PMID 21701511.
- ^ Guainazzi, Angelo; Schärer, Orlando D. (1. 11. 2010). "Using synthetic DNA interstrand crosslinks to elucidate repair pathways and identify new therapeutic targets for cancer chemotherapy". Cellular and Molecular Life Sciences (jezik: engleski). 67 (21): 3683–3697. doi:10.1007/s00018-010-0492-6. ISSN 1420-682X. PMC 3732395. PMID 20730555.
- ^ Cancer, Cleveland Clinic. "Nitrogen Mustard – Chemotherapy Drugs – Chemocare". chemocare.com (jezik: engleski). Arhivirano s originala, 29. 9. 2017. Pristupljeno 9. 10. 2017.
- ^ Jamieson, E. R.; Lippard, S. J. (8. 9. 1999). "Structure, Recognition, and Processing of Cisplatin-DNA Adducts". Chemical Reviews. 99 (9): 2467–2498. doi:10.1021/cr980421n. ISSN 1520-6890. PMID 11749487.
- ^ Poklar N, Pilch DS, Lippard SJ, Redding EA, Dunham SU, Breslauer KJ (juli 1996). "Influence of cisplatin intrastrand crosslinking on the conformation, thermal stability, and energetics of a 20-mer DNA duplex". Proc. Natl. Acad. Sci. U.S.A. 93 (15): 7606–11. Bibcode:1996PNAS...93.7606P. doi:10.1073/pnas.93.15.7606. PMC 38793. PMID 8755522.
- ^ Rudd GN, Hartley JA, Souhami RL (1995). "Persistence of cisplatin-induced DNA interstrand crosslinking in peripheral blood mononuclear cells from elderly and young individuals". Cancer Chemother. Pharmacol. 35 (4): 323–6. doi:10.1007/BF00689452. PMID 7828275. S2CID 24036376.
- ^ Coste, F.; Malinge, J. M.; Serre, L.; Shepard, W.; Roth, M.; Leng, M.; Zelwer, C. (15. 4. 1999). "Crystal structure of a double-stranded DNA containing a cisplatin interstrand cross-link at 1.63 A resolution: hydration at the platinated site". Nucleic Acids Research. 27 (8): 1837–1846. doi:10.1093/nar/27.8.1837. ISSN 0305-1048. PMC 148391. PMID 10101191.
- ^ "Cisplatin". National Cancer Institute (jezik: engleski). 2. 3. 2007. Pristupljeno 9. 10. 2017.
- ^ Cimino, G. D.; Gamper, H. B.; Isaacs, S. T.; Hearst, J. E. (1985). "Psoralens as photoactive probes of nucleic acid structure and function: organic chemistry, photochemistry, and biochemistry". Annual Review of Biochemistry. 54: 1151–1193. doi:10.1146/annurev.bi.54.070185.005443. ISSN 0066-4154. PMID 2411210.
- ^ Qi Wu, Laura A Christensen, Randy J Legerski & Karen M Vasquez, Mismatch repair participates in error-free processing of DNA interstrand crosslinks in human cells,EMBO Reports 6, 6, 551–557 (2005).
- ^ Kirchner, James J.; Sigurdsson, Snorri T.; Hopkins, Paul B. (1. 5. 1992). "Interstrand cross-linking of duplex DNA by nitrous acid: covalent structure of the dG-to-dG cross-link at the sequence 5'-CG". Journal of the American Chemical Society. 114 (11): 4021–4027. doi:10.1021/ja00037a001. ISSN 0002-7863.
- ^ Stone, Michael P.; Cho, Young-Jin; Huang, Hai; Kim, Hye-Young; Kozekov, Ivan D.; Kozekova, Albena; Wang, Hao; Minko, Irina G.; Lloyd, R. Stephen (1. 7. 2008). "Interstrand DNA Cross-Links Induced by α,β-Unsaturated Aldehydes Derived from Lipid Peroxidation and Environmental Sources". Accounts of Chemical Research. 41 (7): 793–804. doi:10.1021/ar700246x. ISSN 0001-4842. PMC 2785109. PMID 18500830.
- ^ Niedernhofer, Laura J.; Daniels, J. Scott; Rouzer, Carol A.; Greene, Rachel E.; Marnett, Lawrence J. (15. 8. 2003). "Malondialdehyde, a Product of Lipid Peroxidation, Is Mutagenic in Human Cells". Journal of Biological Chemistry (jezik: engleski). 278 (33): 31426–31433. doi:10.1074/jbc.m212549200. ISSN 0021-9258. PMID 12775726.
- ^ Dooley, Patricia A.; Zhang, Mingzhou; Korbel, Gregory A.; Nechev, Lubomir V.; Harris, Constance M.; Stone, Michael P.; Harris, Thomas M. (8. 1. 2003). "NMR determination of the conformation of a trimethylene interstrand cross-link in an oligodeoxynucleotide duplex containing a 5'-d(GpC) motif". Journal of the American Chemical Society. 125 (1): 62–72. doi:10.1021/ja0207798. ISSN 0002-7863. PMID 12515507.
- ^ LC Colis; P Raychaudhury; AK Basu (2008). "Mutational specificity of gamma-radiation-induced guanine-thymine and thymine-guanine intrastrand cross-links in mammalian cells and translesion synthesis past the guanine-thymine lesion by human DNA polymerase eta". Biochemistry. 47 (6): 8070–8079. doi:10.1021/bi800529f. PMC 2646719. PMID 18616294.
- ^ Box, Harold C.; Budzinski, Edwin E.; Dawidzik, Jean D.; Wallace, John C.; Evans, Marianne S.; Gobey, Jason S. (1996). "Radiation-Induced Formation of a Crosslink between Base Moieties of Deoxyguanosine and Thymidine in Deoxygenated Solutions of d(CpGpTpA)". Radiation Research. 145 (5): 641–643. Bibcode:1996RadR..145..641B. doi:10.2307/3579285. JSTOR 3579285. PMID 8619032.
- ^ Klein Douwel, Daisy; Boonen, Rick A.C.M.; Long, David T.; Szypowska, Anna A.; Räschle, Markus; Walter, Johannes C.; Knipscheer, Puck (2014). "XPF-ERCC1 Acts in Unhooking DNA Interstrand Crosslinks in Cooperation with FANCD2 and FANCP/SLX4". Molecular Cell. 54 (3): 460–471. doi:10.1016/j.molcel.2014.03.015. PMC 5067070. PMID 24726325.
- ^ DNA repair and mutagenesis. Friedberg, Errol C., Friedberg, Errol C. (2nd izd.). Washington, D.C.: ASM Press. 2006. ISBN 9781555813192. OCLC 59360087.CS1 održavanje: others (link)
- ^ Admiraal, Suzanne J.; O’Brien, Patrick J. (10. 3. 2015). "Base Excision Repair Enzymes Protect Abasic Sites in Duplex DNA from Interstrand Cross-Links". Biochemistry. 54 (9): 1849–1857. doi:10.1021/bi501491z. ISSN 0006-2960. PMC 4404639. PMID 25679877.
- ^ Zhao, Linlin; Xu, Wenyan (2. 12. 2015). "Mutagenic Bypass of an Oxidized Abasic Lesion-Induced DNA Interstrand Cross-Link Analogue by Human Translesion Synthesis DNA Polymerases". Biochemistry. 54 (50): 7409–7422. doi:10.1021/acs.biochem.5b01027. PMC 4700817. PMID 26626537.
- ^ Zhao, Linlin; Xu, Wenyan (21. 10. 2016). "O6-2′-Deoxyguanosine-butylene-O6-2′-deoxyguanosine DNA Interstrand Cross-Links Are Replication-Blocking and Mutagenic DNA Lesions". Chem. Res. Toxicol. 29 (11): 1872–1882. doi:10.1021/acs.chemrestox.6b00278. PMC 5665164. PMID 27768841.
- ^ Cole RS, Levitan D, Sinden RR (1976). "Removal of psoralen interstrand cross-links from DNA of Escherichia coli: mechanism and genetic control". J. Mol. Biol. 103 (1): 39–59. doi:10.1016/0022-2836(76)90051-6. PMID 785009.
- ^ Sinden RR, Cole RS (1978). "Repair of cross-linked DNA and survival of Escherichia coli treated with psoralen and light: effects of mutations influencing genetic recombination and DNA metabolism". J. Bacteriol. 136 (2): 538–47. doi:10.1128/jb.136.2.538-547.1978. PMC 218577. PMID 361714.
- ^ Noll DM, Mason TM, Miller PS (2006). "Formation and repair of interstrand cross-links in DNA". Chem. Rev. 106 (2): 277–301. doi:10.1021/cr040478b. PMC 2505341. PMID 16464006.
- ^ Tretyakova, Natalia; Groehler, Arnold; Ji, Shaofei (2015). "DNA-Protein Cross-links: Formation, Structural Identities, and Biological Outcomes". Acc Chem Res. 48 (6): 1631–44. doi:10.1021/acs.accounts.5b00056. PMC 4704791. PMID 26032357.
- ^ Pendergrast, P.; Chen, Yan; Ebright, Yon; Ebright, Richard (1992). "Determination of the orientationof a DNA binding motif in a protein-DNA complex by photocrosslinking" (PDF). Proc. Natl. Acad. Sci. U.S.A. 89 (21): 10287–10291. Bibcode:1992PNAS...8910287P. doi:10.1073/pnas.89.21.10287. PMC 50323. PMID 1332042. Arhivirano s originala (PDF), 22. 2. 2016. Pristupljeno 17. 3. 2022.
- ^ Smith, Kendric; Martin, Shetlar. "DNA-PROTEIN CROSSLINKS". journal zahtijeva
|journal=(pomoć) - ^ Molina JR, Yang P, Cassivi SD, Schild SE, Adjei AA (2008). "Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship". Mayo Clin. Proc. 83 (5): 584–94. doi:10.4065/83.5.584. PMC 2718421. PMID 18452692.
- ^ a b Taron M, Rosell R, Felip E, Mendez P, Souglakos J, Ronco MS, Queralt C, Majo J, Sanchez JM, Sanchez JJ, Maestre J (2004). "BRCA1 mRNA expression levels as an indicator of chemoresistance in lung cancer". Hum. Mol. Genet. 13 (20): 2443–9. doi:10.1093/hmg/ddh260. PMID 15317748.
- ^ a b Papadaki C, Sfakianaki M, Ioannidis G, Lagoudaki E, Trypaki M, Tryfonidis K, Mavroudis D, Stathopoulos E, Georgoulias V, Souglakos J (2012). "ERCC1 and BRAC1 mRNA expression levels in the primary tumor could predict the effectiveness of the second-line cisplatin-based chemotherapy in pretreated patients with metastatic non-small cell lung cancer". J Thorac Oncol. 7 (4): 663–71. doi:10.1097/JTO.0b013e318244bdd4. PMID 22425915.
- ^ Stingele, Julian; Bellelli, Roberto; Boulton, Simon (septembar 2017). "Mechanisms of DNA–protein crosslink repair". Nature Reviews Molecular Cell Biology. 18 (9): 563–573. doi:10.1038/nrm.2017.56. PMID 28655905. S2CID 9938335.
Vanjski linkovi
uredi- PDB 1AIO – Interaktivna struktura za formiranje cisplatina i DNK adukta
- PDB 204D – Interaktivna struktura za psoralen i umreženu DNK
- Psoralen Ultraviolet A Light Treatment