Excavata
Excavata su jedna od velikih grupa jednoćelijskih protista, uključujući organizme koji žive slobodno, simbionte i neke važne parazite kod ljudi.[1][2][3]
| Excavata (Natkoljeno eukariota, Vremenski raspon: Neoproterozoik → Sadašnjost | |
|---|---|
 Giardia lamblia, parazitska diplomonada | |
| Sistematika | |
| Carstvo | Protista |
| Natkoljeno | Discoba |
| |

Vrh: Jakobida:
1 – Ćelijsko jedro,
2 – Prednji bič,
3 – Ventralni/zadnji bič,
4 – Ventralna brazda za hranu
Sredina - Euglenozoa:
1 – Jedro,
2 – Flagelni džep/rezervoar,
3 – Dorzalni /prednji bič,
4 – Ventralni/zadnji bič,
5 – Citostom/"usni" aparat
Dno - Metamonada
1 – Prednji bič,
2 – parabazno tijelo,
3 – Talasasta (undulatna) membrana,
4 – Zadnji bič,
5 – Jedro,
6 – Aksostil.

1 – Bičevi u apikalnom džepu,
2 – Teka,
3 – Citoplazmatska membrana,
4 – Očna mrlja,
5 – Prednji bič
6 – Ćelijsko jedro,
7 – Hloroplasti.
Procjenjuje se da danas ima 2.300 vrsta Excavata.[4] Većina je heterotrofnih, imaju dvije, četiri ili više bičeva i karakterizira ih ventralni utor za ishranu, koji se koristi za hvatanje i gutanje sitnih čestica uz pomoć struja koje generiraju bičevi. Taj ventralni žlijeb je ono što grupi daje ime. Jedna od grupa uključuje fotosintetske organizme, koji su hloroplasta dobilii u sekundarnoj edosimbiozi sa zelenim algama. Među parazitima u ovoj grupi su Trypanosoma, koji uzrokuju bolest spavanja i Chagovu bolest.
Opis
urediMorfologija ekskavata je vrlo raznolika. Mnogi su okarakterizirani po ventralnom žlijebu za uzimanje hrane, sa karakterističnom ultrastrukturom, podržanom mikrotubulama.[5] Taj ventralni probavni sistem, koji imaju mnogi njihovi predstavnici, iako ne svi, koristi se za hvatanje i gutanje sitnih čestica uz pomoć struje koju generiraju flagele. Suprotno tome, Euglenozoa imaju apikalni ili subapski džep ili invaginaciju umjesto ventralnog žlijeba. On se može sastojati od uskog kanala i rezervoara, koji nisu uključeni u unos hrane, ali heterotrofni oblici imaju citosom, uređaj namijenjen za gutanje čestica, blizak i neovisan o prethodnoj invaginciji. U nekoliko drugih skupina nedostaje udubljenje za uvlačenje, ali klasificirane su s ekskavatima jer imaju sličnu morfologiju ili na osnovu filogenetskih studija.
Excavata su uglavnom heterotrofi (fagofrofi ili osmotrofi). Neki žive slobodno, hrane se česticama, bakterijama ili drugim malim organizmima. Drugi su simbionti u probavnom traktu insekata, pomažu im u probavi celuloze, dok su drugi komensali ili paraziti. Uključene su i neke vrste koje kod ljudi izazivaju ozbiljne bolesti, poput rodova Trypanosoma, Trichomonas i 'Naegleria' '.
Neke Euglenozoa imaju hloroplaste koji su omotani u tri membrane i pigmentirani hlorofilima a i b , sličnim onima kod biljaka, što sugerira da su dobijenene iz simbioze sa zelenim agama. Obično su izgubili citosom, a imaju druge prilagodbe autotrofnom životu, kao što su očne mrlje osjetljive na svjetlost.
Još jedno karakteristično obilježje grupe je organizacija flagelnog aparata, koji obično uključuje dvije, četiri ili više bičeva.[2] Međutim, drugi su ameboidi, a neke grupe imaju ameboidne, amebo-flagelirane i cistne oblike, koje se mogu naizmenično smjenjivati tokom cijelog životnog ciklusa. Tu je i grupa pseudogljiva (Acraside), sa životnim ciklusom koji uključuje faze ameboida, pseudoplasmodija i plodonosnog tijela. Mnoge imaju cjevatre mitohondrijse kriste, koje mogu biti i cjevaste, diskoidni ili, u nekim slučajevima, ravne. Mnogi drugi su anaerobi, budući da žive u okruženju siromašnom kisikom, poput sedimenata ili probavnog trakta nekih životinja. Nedostaju im „klasične“ mitohondrije i često se nazivaju „amitohondrijama“, iako većina, unatoč tome, zadržava mitohondrijsku organelu u izrazito modificiranom obliku (naprimjer, hidrogenosom ili mitosom).
Ekskavate katakterizira bespolno razmnožavanje, binarnom diobom, iako Acrasida formiraju plodno tijelo koja proizvodi spore (slično kao Myxomycota). Seksualna reprodukcija zabilježena je u nekoliko slučajeva, iako u nekim skupinama nije pronađen nijedan primjer, poput Euglenozoa. Sve grupe koje se smatraju kskavatima nemaju ove karakteristike, ali su ovdje uključene na bazi molekulskih analiza.[2] Grupe koje imaju alimentarni žlijeb su: Jakobea, Percolozoa, Fornicata, Trimastix i Malawimonas, iako većina vrsta Percolozoa ima ameboidne vegetativne ćelije. Suprotno tome, Parabasalia i Oxymonadida pretrpjele su transformacije u osnovnom obliku i izgubile su žlijeb za uzimanje hrane. S druge strane, Euglenozoa se hrane preko cjevastog uređaja za gutanje (apikalni ili subapski džep), umjesto ventralnog žlijeba.
Grupe
urediKlasifikacija Excavata je teška. Prema nedavnim filogenetskim analizama, oni su podijeljeni u sedam grupa, koje se mogu smatrati koljenima ili razredima.[3] Rod Malawimonas , mogao bi se također uključiti u ekstavate iako postoje sumnje. Supergrupa Discoba uključuje Jakobida + Tsukubea + Euglenozoa + Percolozoa, dok Metamonada uključuje ostale.
Jakobea
urediJakobea je mala grupa slobodnih, bičastih i heterotrofnih životnih oblika, sa bogatim mitohondrijskim genomom. Imaju dva biča, umetnuta blizu jednog kraja ćelije i povezana sa žlijebom za ishranu. Djelovanje bičeva stvara struju koja pomjera suspendirane čestice prema žlijebu, gdje se razlažu fagocitozom. Ponekad imaju teste (oklop). Uključuje, među ostalim, rodove Jakoba i Reclinomonas .
Tsukubea
urediTsukubea sadrži rod Tsukubamonas , dvobičaste plivačke organizme sa žlijebom za ishranu koji se koristi i za hvatanje plijena.[6]
Euglenozoa
uredi
Euglenozoa su velika skupina bičastih protozoa koja uključuje nekoliko slobodnoplivajućih oblika, kao i nekoliko parazita, od kojih su neki važni patogeni ljudskih bića. Većina ih se hrani gutanjem manjih organizama, obično bakterija ili apsorpcijom. Neki euglenidi poput roda "Euglena" "imaju hloroplaste. Izgubili su citosom i obično imaju druge prilagodbe autotrofnom životu, kao što su očne mrlje, koje su osjetljive na svjetlost. Drugi, poput onih iz roda Trypanosoma uzrokuju ozbiljne bolesti kod ljudi. Većina ima dva biča, obično jedan okrenut prema naprijed, a drugi prema natrag, koji se ubacuju u apikalni ili subapikalni džep.
Percolozoa
urediPercolozoa (= Heterolobosea) je skupina protsta bez hloroplasta, čiji se brojni predstavnici mogu smjestitii između faza ameboida, bičastih oblika i cista. Većina ih živi u tlu, slatkoj vodi ili izmetu. Postoje neki morski i parazitski oblici, uključujući vrstu Naegleria fowleri , koji mogu postati patogeni za ljude i često smrtni.
Metamonada
urediMetamonada je skupina neobičnih flagelata nemaju mitohondrija.[7] Sve Excavata su anaerobi i uglavnom životinjski simbionti. Određene vrste žive u probavnom traktu termita, a imaju važnu ulogu u razgradnji celuloze. Neki oblici su paraziti, uključujući one koji parazitiraju na čovjeku. Tragovi mitohondrije uključuju i hidrogenome koji proizvode vodik. Ćelije mogu imati jedno ili dva jedra. Sve podgrupe imaju bičeve u karakterističnim grupama od po četiri koje su često povezane s jedrima.
- Fornicata: Bičasti amitohondrijski, uglavnom simbiotski ili životinjski paraziti (rodovi Retortamonas, Chilomastix. Giardia, Carpediemonas).
- Parabasalia: Papučaste amitohondrije, obično komensalni organizmi u crijevnom traktu nekih insekata, iako su uključeni i neki patogeni kod ljudi (rod Trichomonas).
- Preaxostyla. Bičasti amitohondrijski, slobodno živeći (Trimastix ) ili žive u crijevima nekih insekata (Oxymonadida).
Geološka starost
urediNajstariji fosilni ostaci koji pripadaju ekskavatima stari su oko 450 miliona godina i odgovaraju rodu Moyeria. Ovaj fosil, koji se nalazi u drevnim slatkovodnim sedimentima, ima jedinstvenu biheliksnu tjelesnu arhitekturu sastavljenu od stopljenih traka veličine 40 do 20 µm. S druge strane, procjena molekularnih satova kalibriranih pomoću najstarijih poznatih fosila svake grupe, procjenjuju da su se Excavata pojavile prije oko 1510-1699 miliona godina.
Filogenija
urediSamo odnos između Euglenozoa i Percolozoa temelji se na robusnim molekulskim podacima; obje grupe imaju i mitohondrije sa diskoidnim grebenima. Većina Jacobea ima cjevaste grebene, kao i većina drugih protista, dok su Metamonada neuobičajene po tome što su izgubile klasične mitohondrije; umjesto toga imaju hidrogenosome, mitosome ili nekarakterizirane organele. Takson, Loukozoa, kontinuirano je redefiniran tako da uključuje širok raspon grupa (uključujući Jacobea i Malawimonas).[8][9]
| Excavata |
| ||||||||||||||||||||||||||||||||||||
Excavata kao parafiletska grupa
urediExcavata su smatrani najprimitivnijim Eukaryaama, uglavnom zato što nekoliko njih nema mitohondrija i zato što se u filogenetskim stablima pojavljuju kao bazni.[10] Ovo sugerira da su ekskavate parafilski taksoni koji uključuju pretke ostalih eukariota. Međutim, pokazano je da je nedostatak mitohondrija u nekim od ovih skupina uzrokovan ranijim gubitkom i da bi početni položaj tih skupina mogao biti posljedica artefakta iz dugog kladusa. Iako su odnosi između različitih skupina još uvijek neizvjesni, neke studije odbacuju njihovu monofiliju i sugeriraju da bi Excavata bila bazna parafiletska eukariotska skupina.[11][12][13][14][15][16][17][18][19][20]
Grupe Excavata prikazane su podebljanim slovima (bold):
| Eukaryota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Drugo istraživanje na aminokiselinama, otkrilo je da su neke grupe više povezane s drugim skupinama eukariota nego sa ostalim ekskavatima; u tom slučaju Diskoba bi bila povezana sa kortikadima, dok bi Metamonada, Ancyromonadida i Malawimonadea bile bliže grupi Podiata:
| Eukaryota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Položaj Excavata kao bazne parafitske eukariotske grupe podržava većina istraživača; međutim, rješenje filogenije Excavata bilo je vrlo izazovno. S druge strane, utvrđeno je da je uključivanje grupe Ancyromonadide u buduće filogenetske studije važno za rješavanje odnosa između svih tih skupina.[21]
Galerija
uredi-

-
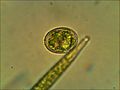 [[Cista kod Trachelomonas (Euglenozoa)
[[Cista kod Trachelomonas (Euglenozoa) -

-

-

-

-

-
 Diferentes etapas (flagelado, ameba, quiste) de Naegleria (Percolozoa)
Diferentes etapas (flagelado, ameba, quiste) de Naegleria (Percolozoa) -

-



.jpg)




.png)


Također pogledajte
urediReference
uredi- ^ Cavalier-Smith, T. 2002. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa Arhivirano 18. 9. 2015. na Wayback Machine. International Journal of Systematic and Evolutionary Microbiology. 52: 297-354.
- ^ a b c Simpson, A.G.B. (2003). Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata[mrtav link]. International Journal of Systematic and Evolutionary Microbiology. 53: 1759-1777.
- ^ a b Adl et al. 2012. The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ^ Adl et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ^ Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- ^ Kamikawa, R., Kolisko, M., Nishimura, Y., Yabuki, A., Brown, M. W., Ishikawa, S. A., ... & Inagaki, Y. (2014). Gene content evolution in discobid mitochondria deduced from the phylogenetic position and complete mitochondrial genome of Tsukubamonas globosa. Genome biology and evolution, 6(2), 306-315.
- ^ Cavalier-Smith T. "The excavate protozoan phyla Metamonada Grassé emend. (Anaeromonadea, Parabasalia, Carpediemonas, Eopharyngia) and Loukozoa emend. (Jakobea, Malawimonas): their evolutionary affinities and new higher taxa". Int. J. Syst. Evol. Microbiol. 53 (Pt 6): 1741–58. PMID 14657102. Arhivirano s originala, 12. 6. 2018. Pristupljeno 12. 2. 2020.
- ^ Parfrey, L.W., Barbero, E., Lasser, E., Dunthorn, M., Bhattacharya, D., Patterson, D.J., Katz, L.A.: Support for the Current Classification of Eukaryotic Diversity http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1713255 Arhivirano 12. 9. 2019. na Wayback Machine
- ^ Hampl, V., Hug, L., Leigh, J. W., Dacks, J. B., Lang, B. F., Simpson, A. G., & Roger, A. J. (2009). Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic “supergroups”. Arhivirano 12. 2. 2020. na Wayback Machine Proceedings of the National Academy of Sciences, 106(10), 3859-3864.
- ^ Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (2006). "Toward automatic reconstruction of a highly resolved tree of life". Science. 311 (5765): 1283–7. PMID 16513982.CS1 održavanje: upotreba parametra authors (link)
- ^ Thomas Cavalier-Smith et al. 2015, Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista. Molecular Phylogenetics and Evolution Volume 93, December 2015, Pages 331–362
- ^ Cavalier-Smith, Thomas; Chao, Ema E.; Lewis, Rhodri (1. 6. 2016). "187-gene phylogeny of protozoan phylum Amoebozoa reveals a new class (Cutosea) of deep-branching, ultrastructurally unique, enveloped marine Lobosa and clarifies amoeba evolution". Molecular Phylogenetics and Evolution. 99: 275–296. doi:10.1016/j.ympev.2016.03.023. PMID 27001604.
- ^ Derelle, Romain; Torruella, Guifré; Klimeš, Vladimír; Brinkmann, Henner; Kim, Eunsoo; Vlček, Čestmír; Lang, B. Franz; Eliáš, Marek (17. 2. 2015). "Bacterial proteins pinpoint a single eukaryotic root". Proceedings of the National Academy of Sciences (jezik: engleski). 112 (7): E693–E699. Bibcode:2015PNAS..112E.693D. doi:10.1073/pnas.1420657112. ISSN 0027-8424. PMC 4343179. PMID 25646484.
- ^ Cavalier-Smith, T.; Chao, E. E.; Snell, E. A.; Berney, C.; Fiore-Donno, A. M.; Lewis, R. (2014). "Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa". Molecular Phylogenetics & Evolution. 81: 71–85. doi:10.1016/j.ympev.2014.08.012. PMID 25152275.
- ^ Cavalier-Smith, Thomas (23. 6. 2010). "Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree". Biology Letters (jezik: engleski). 6 (3): 342–345. doi:10.1098/rsbl.2009.0948. ISSN 1744-9561. PMC 2880060. PMID 20031978.
- ^ He, Ding; Fiz-Palacios, Omar; Fu, Cheng-Jie; Fehling, Johanna; Tsai, Chun-Chieh; Baldauf, Sandra L. (2014). "An Alternative Root for the Eukaryote Tree of Life". Current Biology. 24 (4): 465–470. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2014.01.036. PMID 24508168.
- ^ Cavelier Smith (2013). "Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa". European Journal of Protistology. 49 (2): 115–178. doi:10.1016/j.ejop.2012.06.001. PMID 23085100.
- ^ Hug, Laura A.; Baker, Brett J.; Anantharaman, Karthik; Brown, Christopher T.; Probst, Alexander J.; Castelle, Cindy J.; Butterfield, Cristina N.; Hernsdorf, Alex W.; Amano, Yuki (11. 4. 2016). "A new view of the tree of life". Nature Microbiology (jezik: engleski). 1 (5): 16048. doi:10.1038/nmicrobiol.2016.48. ISSN 2058-5276. PMID 27572647.
- ^ Cavalier-Smith, Thomas (5. 9. 2017). "Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences". Protoplasma (jezik: engleski). 255 (1): 297–357. doi:10.1007/s00709-017-1147-3. ISSN 0033-183X. PMC 5756292. PMID 28875267.
- ^ Cavalier-Smith, Thomas (2017). "Euglenoid pellicle morphogenesis and evolution in light of comparative ultrastructure and trypanosomatid biology: semi-conservative microtubule/strip duplication, strip shaping and transformation". European Journal of Protistology. 61 (Pt A): 137–179. doi:10.1016/j.ejop.2017.09.002. PMID 29073503.
- ^ Brown, M. W., Heiss, A. A., Kamikawa, R., Inagaki, Y., Yabuki, A., Tice, A. K., ... & Roger, A. J. (2018). Phylogenomics places orphan protistan lineages in a novel eukaryotic super-group. Genome biology and evolution, 10(2), 427-433.