Pužnica
Pužnica ili puž je dio unutrašnjeg uha koji je uključen u čulo sluha. To je šupljina u obliku spirale u koštanom labirintu, kod ljudi, koji ima 2,75 zavoja oko svoje ose, zvane modiolus.[1][2] Ključna komponenta pužnice je Cortijev organ, senzorni organ sluha, koji je raspoređen duž pregrade koja razdvaja komore za tečnost u zavojitoj suženoj cijevi pužnice.
| Pužnica | |
|---|---|
 Poprečni presjek pužnice | |
| Detalji | |
| Dio | Unutrašnje uho |
| Sistem | Čulo sluha |
| Identifikatori | |
| Latinski | 'Cochlea' |
| MeSH | D003051 |
| NeuroLex ID | birnlex_1190 |
| TA98 | A15.3.03.025 |
| TA2 | 6964 |
| FMA | 60201 |
| Anatomska terminologija | |

Latinski naziv pužnice kohlea potiče iz starogrčkog κοχλίας – kokhlias = spirala, puževa ljuska).
Struktura
uredi
Pužnica je spiralna , šuplja, konusna komora kostiju, u kojoj se valovi šire od "baze" (blizu srednjeg uha i ovalnog okna) do "vrha" (gornjeg dijela ili središta spirale). Spiralni kanal pužnice dio je koštanog labirinta unutrašnjeg uha, dug oko 30 mm i koji čini 2¾ zavoja oko modiolusa. Njene strukture uključuju:
- Tri skale ili komore:
- vestibulski kanal ili scala vestibuli (koji sadrži periilimfu), koji leži superiornije od pužnog kanala i naslanja se na ovalno okno
- timpanijski kanal ili scala tympani (koji sadrži perilimfu) i leži ispod pužnog kanala i završava na okruglom oknu
- pućnični kanal ili scala media (koji sadrži endolimfu), područje visoke koncentracije kalijevih iona u koje se projiciraju ćelijske stereocilije, ćelijske treplje
- Pužnjični trijem, mjesto spajanja timpanijskog i vestibulskog kanala, na vrhu pužnice
- Reissnerova membrana, koja odvaja vestibulskii kanal od pužnog kanala
- Koštano-spiralna lamina, glavni strukturni element koji razdvaja pužni kanal od bubnjića
- Bazilarna membrana, glavni strukturni element koji odvaja pužnjičnii kanal od bubnjića i određuje svojstva mehaničkog širenja talasa pužneve pregrade
- Cortijev organ, senzorni epitel, ćelijski sloj na bazilarnoj membrani, u kojem se treplje senzornih ćelija napajaju potencijalnom razlikom između perilimfe i endolimfe
- ćelije treplje, senzorne ćelije u Cortijevom organu, prekrivene strukturama nalik dlakama zvanim stereocilije ili stereotreplje
- Spiralni ligament.
Pužnica je dio unutrašnjeg uha koji izgleda poput puževe ljuske (puž je grčki naziv za puža).[3] Pužnica prima zvuk u obliku vibracije koje uzrokuju kretanje stereocilija. Stereocilije zatim pretvaraju ove vibracije u nervni impuls koji se prenose u mozak da bi se protumačili. Dva od tri dijela tekućina su kanali, a treći je osjetljivi 'Cortijev organ' koji otkriva impulsni pritisak koji putuje dužim živcem do mozga. Dva kanala nazivaju se vestibulskim i timpanijskim kanalom.
Mikroanatomija
urediZidovi šuplje pužnice napravljene su od kostiju, sa tankom, nježnim oblogom od epitelnog tkiva. Ova spirala čiji je prostor podijeljen većim dijelom dužine unutrašnjim membranskim pregradama. Ovom pregradnom membranom nastaju dva vanjska prostora ispunjena tekućinom („kanali“ ili „skale“). Na vrhu cijevi namotanih u obliku puževa, nalazi se preokret u smjeru kretanja tekućina, mijenjajući tako vestibulski u timpanijski kanal. Ovo područje se naziva helikotrijem. Ovaj nastavak na helikotrijemu omogućava da se ovalnim oknom tekućina potiskuje u vestibulski kanal da bi se pomaknula natrag kretanjem u bubnju i otklonom okružnog prozora; budući da je tekućinu gotovo nemoguć stisnuti pod pritiskom, a koštani zidovi su kruti, neophodno je sačuvati količinu tekućinw da negdje izađe.
Uzdužna pregrada koja dijeli veći dio pužnice sama je po sebi cijev punjena tekućinom, treći kanal. Ovaj središnji stub naziva se pužni kanal. Njegova tečnost, endolimfa, takođe sadrži elektrolite i proteine, ali se hemijski prilično razlikuje od perilimfe. Dok je perilimfa bogata natrijevim ionima, endolimfa je bogata kalijevim, što stvara ionski, električni potencijal.
Ćelije sa dlačicama raspoređene su u četiri reda u Cortijevom organu, duž cijele dužine pužnice. Tri se reda sastoje od vanjskih trepljastih ćelija (OHC), jedan crveni od unutrašnjih trepljastih ćelija (IHC). Unutrašnje ćelije sadrže glavne neuronske izlaze pužnice. Umesto toga, vanjske trepljaste ćelije uglavnom "primaju" neuronski ulaz iz mozga, što utiče na njihovu pokretljivost kao dio mehaničkog pretpojačala pužnice. Ulaz u OHC dolazi iz masnog tijela, putem medijalnog olivopužničnog snopa.
Pužni kanal je sam po sebi gotovo jednako složen kao i samo uho. Pužovod je na tri strane omeđen bazilarnom membranom, stria vascularis i Reissnerovom membranom. Stria vascularis je bogati sloj kapilarima i sekretornim ćelijama; Reissnerova membrana je tanka membrana koja razdvaja endolimfu od perilimfe; bazilarna membrana je mehanički pomalo kruta membrana koja podupire receptorski organ za sluh, Cortijev organ i određuje svojstva mehaničkog širenja talasa u pužnom sistemu.
Funkcija
urediPužnica je ispunjena vodenastom tečnošću, zvanom endolimfa, koja se kreće kao odgovor na vibracije koje dolaze iz srednjeg uha kroz ovalni prozor. Kako se tekućina kreće, ppuže i kohlearna pregrada (bazilarna membrana i Cortijev organ); hiljade trepljastih ćelija osjete kretanje putem svojih stereocilija i pretvaraju to kretanje u električne signale koji se putem neurotransmitera prenose na hiljade nervnih ćelija. Ovi primarni slušni neuroni transformiraju signale u elektrohemijske impulse poznate kao akcijski potencijal, koji putuju duž slušnog živca do struktura u moždanom stablu radi dalje obrade.
Slušanje
urediKost skeleta srednjeg uha stapes (stremen) prenosi vibracije na fenestra ovalis (ovalno okno) na vanjskoj strani pužnice, koja vibrira perilimfu u vestibulskii kanal (gornja komora pužnice). Koščice su ključne za efikasno spajanje zvučnih talasa u pužnici, jer je okolina sistem puža tečna membrana i potreban je veći pritisak da se zvuk prenosi kroz talase fluid-membrana nego za zrak. Povećanje pritiska postiže se smanjenjem odnosa površine od bubne opne (bubanj) do ovalnog okna (stremenska kost) za 20 x. Kako je pritisak = sila / površina, rezultira porastom pritiska od oko 20 puta od prvobitnog pritiska zvučnog talasa u zrakuu. Ovo pojačanje je oblik podudaranja impedencije – da se zvučni talas koji putuje kroz zrak podudara sa onim koji putuje u sistemu fluid-membrana.
Na dnu pužnice, svaki kanal završava se membranskim portalom, koji je okrenut ka šupljini srednjeg uha: vestibulski kanal završava na ovalnom oknu, gdje se nalazi koščica stapes (stremen). Ona vibrira kada se pritisak prenosi preko osikulskog lanca. Talas u perilimfi odmiče se od podnožja prema helikotrrijemu. Budući da ti fluidni rtalasii pomiču pužničnuu pregradu koja razdvaja kanale gore-dolje, talasi imaju odgovarajući simetrični dio u perilimfi "bubnjića", koji završava na okruglom oknu i ispupčen je kad se izboči ovalno okno.
Perilimfa u vestibulskm kanalu i endolimfa u pužnom kanalu djeluju mehanički kao jedan kanal, a razdvaja ih samo vrlo tanka Reissnerova membrana.
Vibracije endolimfe u pužnom kanalu istiskuju bazilarnu membranu po obrascu koji doseže udaljenost od ovalnog okna, ovisno o frekvenciji zvučnog talasa. Cortijev organ vibrira zbog vanjskih trepljastih ćelija, što dodatno pojačava ove vibracije. Unutzrašnje treplje se tada istiskuju vibracijama u tečnosti, a depolariziraju se prilivom K+, preko njihovih vršnih veza, povezanih kanala i šalju svoje signale putem neurotransmitera do primarnih slušnih neurona spiralne ganglije.
Trepljaste ćelije u Cortijevom organu podešene su na određene frekvencije zvuka zbog svog smještaja u pužnici, stupnja ukočenosti bazilarne membrane.[4] Ova krutost je, između ostalog, posljedica debljine i širine bazilarne membrane,[5] koja je po dužini pužnice najtvrđa najbliže svom početku na ovalnom oknu, gdje stipes vodi vibracije koje dolaze iz bubnjića. Budući da je tamo njegova ukočenost velika, omogućava samo vibracijama visoke frekvencije da pokreću bazilarnu membranu, a time i ćelijske treplje. Što talas dalje putuje prema vrhu pužnice ("helikotrijema"), bazilarna membrana je manje ukočena; tako niže frekvencije putuju niz cijev, a manje krutu membranu oni najlakše premještaju tamo gdje to smanjena krutost dopušta: to jest, kako je bazilarna membrana sve manje i manje kruta, talasi se usporavaju i ona bolje reagira na niže frekvencije. Pored toga, kod sisara se pužnica namotava, gdje je pokazano da pojačava niskofrekventne vibracije dok putuju kroz zavojnicu ispunjenu tečnošću.[6] Ovaj prostorni raspored prijema zvuka naziva se tonotopija.
Za vrlo niske frekvencije (ispod 20 Hz), talasi se šire cijelom trasom pužnice - diferencijalno prema vestibularnom kanalu i timpanijskom kanalu sve do helikotreme '. Ovako niske frekvencije još uvijek aktiviraju Cortijev organ u određenoj mjeri, ali su preniske da bi izazvale percepciju visina. Veće frekvencije se ne šire u "helikotremu" zbog tonotopije posredovane krutošću.
Vrlo snažno kretanje bazilarne membrane zbog vrlo glasne buke može uzrokovati odumiranje trepljastih ćelija. Ovo je čest uzrok djelimičnog gubitka sluha i razlog zašto korisnici vatrenog oružja ili teške mehanizacije često nose naušnik ili čep za uši.
Pojačanje trepljastih ćelija
urediPužnica ne samo da „prima“ zvuk, već zdrava pužnica „generira“ i pojačava zvuk kada je to potrebno. Tamo gdje je organizmu potreban mehanizam da čuje vrlo slabe zvukove, pužnica se pojačava obrnutim transdukcijama OHC-a, pretvarajući električne signale natrag u mehaničke, u konfiguraciji pozitivne povratne sprege. OHC imaju na svojim vanjskim membranama proteinski motor zvan prestin; generira dodatno kretanje koje se spaja natrag sa talasom fluid-membrana. Ovo "aktivno pojačalo" je neophodno u sposobnosti uha da pojačava slabe zvukove.[7][8]
Aktivno pojačalo takođe dovodi do pojave vibracija zvučnog talasa koje se iz pužnice emitiraju natrag u ušni kanal kroz srednje uho (otoakustičke emisije).
Otoakustičke emisije
urediOtoakustična emisija nastaje uslijed talasa koji izlazi iz pužnice kroz ovalno okno i širi se natrag kroz srednje uho do bubne opne i kroz ušni kanal, gdje ga mikrofon može podići. Otoakustične emisije su važne u nekim vrstama testova za oštećenje sluha, jer su prisutne kada pužnica dobro funkcionira, a manje kada pati od gubitka aktivnosti OHC.
Uloga međućelijskih veza
urediProteini između spoja, nazvani koneksini, u pužnici imaju važnu ulogu u funkcioniranju sluha.[9] Utvrđeno je da mutacije gena za spajanje ćelija uzrokuju sindromsku i nesindromsku gluhoću.[10] Određeni koneksini, uključujući koneksin 30 i koneksin 26, rasprostranjeni su u dva različita sistema uskih međućelijskih veza, koje se nalaze u pužnici. Mreža za spajanje epitelnih ćelija spaja nesenzorne epitelne ćelije, dok mreža za spajanje vezivnog tkiva spaja ćelije tog tkiva. Kanali za spajanje kalijevih iona vraćaju kalij nazad u endolimfu, nakon mehanotransdukcije u trepljaste ćelije.[11] Određeni koneksini, uključujući koneksin 30 i koneksin 26, rasprostranjeni su u dva različita sistema spoja jaza koji se nalaze u pužnici. Mreža za spajanje epitelnih ćelija spaja nesenzorne epitelne ćelije, dok mreža za spajanje vezivnog tkiva spaja ćelije vezivnog tkiva. Kanali za spajanje kalijuma joni kalijuma vraćaju se nazad u endolimfu nakon mehanotransdukcije u trepljaste ćelije.[12]
Dodatne slike
uredi-
 Desni koštani labirint. Bočni pogled.
Desni koštani labirint. Bočni pogled. -
 Donji koštani labirint.
Donji koštani labirint. -
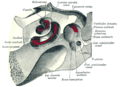 Vestibulum i pužnica, odozgo
Vestibulum i pužnica, odozgo -
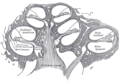 Poprečni presjek pužnice
Poprečni presjek pužnice




Također pogledajte
urediReference
uredi- ^ Anne M. Gilroy; Brian R. MacPherson; Lawrence M. Ross (2008). Atlas of anatomy. Thieme. str. 536. ISBN 978-1-60406-151-2.
- ^ Moore & Dalley (1999). Clinically Oriented Anatomy (4th izd.). str. 974. ISBN 0-683-06141-0.
- ^ The Kingfisher children's encyclopedia. Kingfisher Publications. (3rd ed., fully rev. and updated izd.). New York: Kingfisher. 2012 [2011]. ISBN 9780753468142. OCLC 796083112.CS1 održavanje: others (link)
- ^ Guenter Ehret (Dec 1978). "Stiffness gradient along the basilar membrane as a way for spatial frequency analysis within the cochlea" (PDF). J Acoust Soc Am. 64 (6): 1723–6. doi:10.1121/1.382153. PMID 739099.
- ^ Camhi, J. Neuroethology: nerve cells and the natural behavior of animals. Sinauer Associates, 1984.
- ^ Manoussaki D, Chadwick RS, Ketten DR, Arruda J, Dimitriadis EK, O'Malley JT (2008). "The influence of cochlear shape on low-frequency hearing". Proc Natl Acad Sci U S A. 105 (16): 6162–6166. Bibcode:2008PNAS..105.6162M. doi:10.1073/pnas.0710037105. PMC 2299218. PMID 18413615.
- ^ Ashmore, Jonathan Felix (1987). "A fast motile response in guinea-pig outer hair cells: the cellular basis of the cochlear amplifier". The Journal of Physiology (jezik: engleski). 388 (1): 323–347. doi:10.1113/jphysiol.1987.sp016617. ISSN 1469-7793. PMC 1192551. PMID 3656195.
- ^ Ashmore, Jonathan (2008). "Cochlear Outer Hair Cell Motility". Physiological Reviews (jezik: engleski). 88 (1): 173–210. doi:10.1152/physrev.00044.2006. ISSN 0031-9333. PMID 18195086. S2CID 17722638.
- ^ Zhao, H. -B.; Kikuchi, T.; Ngezahayo, A.; White, T. W. (2006). "Gap Junctions and Cochlear Homeostasis". Journal of Membrane Biology. 209 (2–3): 177–186. doi:10.1007/s00232-005-0832-x. PMC 1609193. PMID 16773501.
- ^ Erbe, C. B.; Harris, K. C.; Runge-Samuelson, C. L.; Flanary, V. A.; Wackym, P. A. (2004). "Connexin 26 and Connexin 30 Mutations in Children with Nonsyndromic Hearing Loss". The Laryngoscope. 114 (4): 607–611. doi:10.1097/00005537-200404000-00003. PMID 15064611.
- ^ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Takasaka, T.; Adams, J. C. (2000). "Gap junction systems in the mammalian cochlea". Brain Research. Brain Research Reviews. 32 (1): 163–166. doi:10.1016/S0165-0173(99)00076-4. PMID 10751665. S2CID 11292387.
- ^ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Adams, J. C. (1995). "Gap junctions in the rat cochlea: Immunohistochemical and ultrastructural analysis". Anatomy and Embryology. 191 (2): 101–118. doi:10.1007/BF00186783. PMID 7726389.
Dopunska literatura
uredi- Dallos, Peter; Popper, Arthur N.; Fay, Richard R. The Cochlea.
- Imbert, Michel; Kay, R. H. (1992). Audition. ISBN 9780262023313.
- Jahn, Anthony F.; Santos-Sacchi, Joseph (2001). Physiology of the Ear. ISBN 9781565939943.
- Roeser, Ross J.; Valente, Michael; Hosford-Dunn, Holly (2007). Audiology. ISBN 9781588905420.
Vanjski linkovi
uredi- Cochlea na US National Library of Medicine Medical Subject Headings (MeSH)
- "Promenade 'Round the Cochlea" Arhivirano 9. 1. 2002. na Wayback Machine by R. Pujol, S. Blatrix, T. Pujol et al. at University of Montpellier
- "Histology Videos of The Ear"