Polifosfat
Polifosfati su soli ili estri polimernih oksianiona koji su formirani od tetraedarskog PO4 fosfata, strukturne jedinice koja je vezana za zajednički atom kisika. Polifosfati mogu imati linearnu ili prstenastu strukturu. U biološkim procesima i supstancama, polifosfatni estri adenozin-difosfat (ADP) i ATP uključeni su u skladištenje/otpuštanje energije. Mnoštvo polifosfata ima primjenu u mineralnim prećišćavanjima komunalnih voda, uglavnom u količinama 1 do 5 ppm.[1]GTP, CTP i UTP su također važni nukleotidi u sintezi proteina i lipida, kao i metabolizmu ugljikohidrata.
Struktura
uredi-
 Struktura trifosforne kiseline
Struktura trifosforne kiseline -
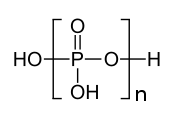
-
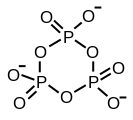 Ciklični trimetafosfat
Ciklični trimetafosfat -
 Adenozin-difosfat (ADP)
Adenozin-difosfat (ADP)




Struktura tripolifosforne kiseline ilustrira načela koja definiraju strukture polifosfata. Sastoji se od tri tetraedne PO4 jedinice povezane tako da ima zajednički kisikov centar. Za linearne lance, fosforna grupa na kraju ima jedan zajednički oksid, a ostali fosfori u centrima dijele dva oksidna centra. Odgovarajući fosfati se pretvaraju u kiseline gubitkom kiselih protona. U slučaju cikličnih trimera, svaki tetraedar dijeli dva čvora sa susjednim tetraedrom.
Postoje tri zajednička moguča ugla. Ovaj motiv predstavlja umreženi linearni polimera. Umreženi polifosfati imaju listnatu strukture filosilikata, ali takve strukture javljaju se samo u ekstremnim uvjetima.
Formiranje i sinteza
urediPolifosfati nastaju polimerizacijom izvedenica fosforne kiseline. Proces počinje sa dvije fosfatne jedinice koje ulaze u zajedničku kondenzacijsku reakciju:
- 2 HPO42− P2O74− + H2O

Kondenzacija je prikazana kao ravnotežna zato što je također moguća i povratna reakcija, tj. hidroliza. Proces se može nastaviti postepeno: u svakom koraku, na lanac se dodaje naredna jedinica PO3, kao što je označeno u zagradama u ilustraciji polifosforne kiseline. P4O10vidi se kao krajnji proizvod kondenzacijskih reakcija, gdje svaki tetraedar ima tri zajednička ugla sa ostalima. Nasuprot tome, složena mješavina polimera nastaje kada se fosfor-pentoksidu dodaje mala količina vode.
Kiselinsko-bazna i kompleksacijska svojstva
urediPolifosfati su slabe baze. Usamljeni par elektrona na atomu kisika može biti doniran u ion vodika (proton) ili metalni ion u tipskim interakcijama Lewisovim kiseline - Lewisove baze. To ima duboko značenje u biologiji. Naprimjer, oko 25% adenozin-trifosfata se protonira u vodenoj otopini pri pH 7.[2]
- ATP4− + H+ ATPH3−, pKa 6.6

Dalje protonacije javljaju se na nižim pH vrijednostima.
"Visokoenegetska" fosfatna veza
urediATP formira helatni komplekse sa metalnim ionima. konstanta stabilnosti za ravnotežu
- ATP4− + Mg2+ MgATP2−, log β 4
posebno je velika.[3] Formiranje kompleksa magnezija je ključni element u procesu hidrolize ATP, jer slabi vezu između terminalne fosfatne grupe i ostatka molekule.[2][4]
Po biološkim standardima, otpuštena energija u hidrolizi ATP,
- ATP4− + H2O → ADP3− + Pi−
na ΔG –36,8 kJ mol−1 je velika. Pi je lanac neorganskog fosfata, koji se protonizira pri biološkom pH. Međutim, po neorganskim standardima, velik je. Termin "visokoenergetski" odnosi se na činjenicu da su relativno energijski bogati u odnosu na iznose energije koja se oslobađa u reakcijama organske hrmija koje se dešavaju u živim sistemima.
Visokopolimerni neorganski polifosfati
urediDobro su poznati polifosfati sa visokom molekulskom masom. Među takvim derivatima su jestaklasta, tj. amorfna Grahamova so. Kristalni polifosfati visoke molekularne težine uključuju Kurrolovu i Maddrellovu so.
Takve vrste imaju formulu [NaPO3]n[NaPO3(OH)]2, gdje n može biti veći od 2000. U smislu njihove strukture, ovi polimeri sastoje se od PO3– "monomera", sa lancima koji se završavaju protoniziranim fosfatima.[5]
Prirodni polifosfati
urediL. Liberman (1890.) u živim organizmima nije pronašao visoko polimerne anorganske polifosfate. Ovi spojevi su linearni polimeri koji sadrže nekoliko do nekoliko stotina ortofosfornih ostataka povezanih energetski bogatim fosfoanhidridnim vezama.
Ranije, ovi spojevi smatrali su se ili "molekularnim fosilima" ili kao samo fosfor i izvor energije za preživljavanje mikroorganizama pod ekstremnim uvjetima. Za njih se sada zna da imaju i regulatorne uloge i da se nalaze u predstavnicima svih carstava živih organizama, gdje učestvuju u metaboličkim korekcijama i kontroli i genetičkih i enzimskih razina. Polifosfati su direktno uključeni u mijenjanje genetičkog programa za obilježja eksponencijalne faze rasta bakterija u programu opstanka ćelije pod stacionarnim uvjetima, "života u sporoj liniji". Oni učestvuju u mnogim regulatornim mehanizama bakterija:
- Učestvuju u indukciji rpoS, RNK-polimerazne podjedinice koja je odgovorna za ekspresiju velike grupe gena uključenih u prilagođavanja stacionarne faze rasta i u mnoge stresne agense.
- Važni su za mobilnu pokretljivost, formiranje biofilma i virulencije.
- Polifosfati i eksopolifosfataza učestvuju u regulaciji nivoa faktora snažnih odgovora, gvanozin 5'-difosfat 3'-difosfata (ppGpp), koji je drugi glasnik u bakterijskim ćelijama.
- Polifosfati učestvuju u formiranju kanala preko žive ćelijske membrane. Navedeni kanali koji formiraju polifosfat i poli-B-hidroksibutirat sa Ca2+ uključeni su u procese transporta u različitim organizmima.
- Važna funkcija polifosfata u mikroorganizmima i prokariotskim — i nižim skim organizmima— pogodnoa je za upravljanje promjenama u okolišnim uvjetima jer obezbjeđuju fosfate i energetske rezerve. Polifosfati su prisutni u životinjskim ćelijama, a postoje i mnogi podaci o njihovom učešću u regulatornim procesima tokom razvoja i mobilnih proliferacija i diferencijacija —posebno u koštanim tkivima i mozgu.
Kod ljudi, pokazalo se da polifosfati imaju ključnu ulogu u koagulaciji krvi, a u produkciji trombocita[6] oni aktiviraju faktor XII koji je neophodna za formiranje krvnih ugrušaka. Osim toga, polifosfati izvedeni iz trombocita aktiviraju faktora XII (Hageman faktor) zgrušavanja krvi koji inicira formiranje fibrina i stvaranje jednog od proinflamatornih posrednika, bradikinina koji doprinosi curenju iz krvnih sudova i tromboza.[7][8]
Neorganski polifosfati imaju ključnu ulogu u toleranciji ćelija kvasca za toksičnost teških metalnih kationa.[9]
Također pogledajte
urediReference
uredi- ^ "Arhivirana kopija" (PDF). Arhivirano s originala (PDF), 19. 4. 2011. Pristupljeno 24. 1. 2017.CS1 održavanje: arhivirana kopija u naslovu (link)
- ^ a b Storer A, Cornish-Bowden A (1976). "Concentration of MgATP2- and other ions in solution. Calculation of the true concentrations of species present in mixtures of associating ions". Biochem J. 159 (1): 1–5. PMC 1164030. PMID 11772.CS1 održavanje: upotreba parametra authors (link)
- ^ Wilson J, Chin A (1991). "Chelation of divalent cations by ATP, studied by titration calorimetry". Anal Biochem. 193 (1): 16–9. doi:10.1016/0003-2697(91)90036-S. PMID 1645933.CS1 održavanje: upotreba parametra authors (link)
- ^ Garfinkel L, Altschuld R, Garfinkel D (1986). "Magnesium in cardiac energy metabolism". J Mol Cell Cardiol. 18 (10): 1003–13. doi:10.1016/S0022-2828(86)80289-9. PMID 3537318.CS1 održavanje: upotreba parametra authors (link)
- ^ Klaus Schrödter, Gerhard Bettermann, Thomas Staffel, Friedrich Wahl, Thomas Klein, Thomas Hofmann "Phosphoric Acid and Phosphates" in Ullmann’s Encyclopedia of Industrial Chemistry 2008, Wiley-VCH, Weinheim. doi:10.1002/14356007.a19_465.pub3
- ^ Ruiz FA, Lea CR, Oldfield E, Docampo R (2004). "Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes". J Biol Chem. 279 (43): 44250–7. doi:10.1074/jbc.M406261200. PMID 15308650.CS1 održavanje: upotreba parametra authors (link)
- ^ Müller F, Mutch NJ, Schenk WA, Smith SA, Esterl L, Spronk HM, Schmidbauer S, Gahl WA, Morrissey JH, Renné T (2009). "Platelet polyphosphates are proinflammatory and procoagulant mediators in vivo". Cell. 139 (6): 1143–56. doi:10.1016/j.cell.2009.11.001. PMC 2796262. PMID 20005807.CS1 održavanje: upotreba parametra authors (link)
- ^ "Newly discovered mechanism by which blood clots form". physorg.com. 10. 12. 2009. Pristupljeno 13. 12. 2009.
- ^ Andreeva N, Ryazanova L, Dmitriev V, Kulakovskaya T, Kulaev I (2013). "Adaptation of Saccharomyces cerevisiae to toxic manganese concentration triggers changes in inorganic polyphosphates". FEMS Yeast Res. 13 (5): 463–470. doi:10.1111/1567-1364.12049. PMID 23663411.CS1 održavanje: upotreba parametra authors (link)
Vanjski linkovi
uredi- Pavlov E, Grimbly C, Diao CT, French RJ (septembar 2005). "A high-conductance mode of a poly-3-hydroxybutyrate/calcium/polyphosphate channel isolated from competent Escherichia coli cells". FEBS Lett. 579 (23): 5187–92. doi:10.1016/j.febslet.2005.08.032. PMID 16150446.CS1 održavanje: upotreba parametra authors (link)
- Kulaev I, Vagabov V, Kulakovskaya T (1999). "New aspects of inorganic polyphosphate metabolism and function". J. Biosci. Bioeng. 88 (2): 111–29. doi:10.1016/S1389-1723(99)80189-3. PMID 16232585.CS1 održavanje: upotreba parametra authors (link)
- Kulaev I, Kulakovskaya T (2000). "Polyphosphate and phosphate pump". Annu. Rev. Microbiol. 54: 709–34. doi:10.1146/annurev.micro.54.1.709. PMID 11018142.CS1 održavanje: upotreba parametra authors (link)[mrtav link]