Trilobiti
Trilobita ili trilobiti, trilobite (lat. tres lobulos = tri režnja, trorežnjevito, trodijelno) su izumrli razred Arthropoda, u potkoljenu Trilobitomorpha. Oni su najkarakterističniji fosili za paleozojsku eru (ranije "primarno doba"). Opisano ih je gotovo 4.000 vrsta.[1]
| Trilobita Vremenski raspon: Kambrij – Gornji perm | |
|---|---|
 Fosil Phacops rana, trilobita iz devona | |
| Sistematika | |
| Carstvo | Animalia |
| Koljeno | Arthropoda |
| Potkoljeno | Trilobitomorpha † |
| Razred | Trilobita † |
| Agnostida Redlichiida | |



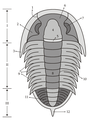
1 – predočni prostor;
2 – palpebralno područje;
3 – zaočni prostor;
4 – posterolateralna projekcija;
5 – okcipitalni prsten;
6 – glabela;
7 – stražnja površina;
8 – bočna granica;
9 – librigenalna površina;
10 – preglabelska površina


.png)


Pojavili su se u kambrijskom periodu (na početku paleozoika, prije oko 540 miliona godina), kada su se pojavili edijakarski organizmi, ali su se počeli diverzificirati u donjem kambriju. Nakon masovnog izumiranja u kasnom kambriju preživjeli su samo oblici koji su naseljavali pelagijske dubokovodne sredine.
Tokom ordovicija dostigli su maksimalnu raznolikost i zauzeli gotovo sve morske ekološke niše. Od silura je bilo malo promjena, sve dok u srednje- i donjedevonskoj krizi nisu pretrpjeli značajno smanjenje, izumiranjem svih redova, osim Proetida . Tokom karbona, predstavici ovih grups su rijetke i ograničeni na grebenove. Posljednji trilobiti, već stanovnici samo plitkih voda, nestali su za vrijeme granične permo-trijaske krize, prije oko 250 miliona godina, zajedno sa Eurypterida ili „morski škorpijama Blastozoa, nekim vrstama riba, spužvi, bodljokožaca itd. Njegovo prisustvo na Zemlji, dakle, trajalo tokom paleozoika, gotovo 300 miliona godina. Fosilni ostaci trilobita toliko su obilni i toliko su proučeni da su vjerovatno najpoznatija skupina izumrlih životinja.
U početku su se smatrali precima rakova (posebno kopnenih Cochinealla vlažnih staništa, koji imaju određene zajedničke karakteristike) ili čak svih člankonožaca (jer su se prvi pojavili u fosilnim zapisima) . Danas se smatraju nezavisnom skupinom, izdvojene od Mandibulata i Chelicerata.
Morfologija uredi
Trilobiti su imali ravno i glatko tijelo, više ili manje ovalno, a podijeljenu u tri tagme, cefalon (glava), grudni koš i pigidij; grudni koš i pigidij formiraju trup. Imali su dvije uzdužne brazde koje tijelo dijele na tri jasno definirana režnja (od čega dolazi i ime): srednji (koji se u zove "glabela" i "trunkus") i dva bočna (tzv. gena ili obrazi u cefalonu i pleura u trupu). Dorzalni tegument bio je gusta kutikula impregnirana kalcij-karbonatom, koji je olakšavao njihovu fosilizaciju. Veličina im je varirala od nekoliko milimetara do više od 60 cm, kod nekih džinovskih vrsta..[2]
-
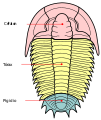 Tagmas
Tagmas -
 Morfología
Morfología -
 Céfalon
Céfalon



Glava uredi
Glava (cephalon) rezultat je potpune fuzije različitih segmenata i ne pokazuje vanjske ostatke metamerizacije. Gēna se proteže sa strana i iza dva genalnog vrha ; gena se šavom dijeli na dva dijela: unutrašnja, fiksna gēna (fiksigena) i vanjska, mobilna gena (librigena) . Struktura koja je formirana od glabele i fiksigena naziva se kranidija. Na ventralnoj strani je hipostom, plak koji je suspendiran ispod glabele, a koji štiti usta.
Na genama postoji par velikih kompozitnih očiju, iznenađujuće evoluiranih (kod nekih vrsta smještenih na stabljikama), analogno onima kod sadašnjih srodnika, kao što su rakovi i insekti. U stvari, trilobiti su bili prve životinje koje su imale složene oči, što je vjerovatno uticalo na njihov evolucijski uspjeh. U ventralnom delu cefalona ubacuju se antene vrlo slične jedne drugima. Antene su, po svom položaju rasporedu, jednake onima kod insekata i Myriapoda i prvom paru kod rakova.
Toraks uredi
Toraks je formiran od između 2 i 40 metamera, dobro je diferenciran; bili su artikulirani jedni s drugima, omogućavajući spajanje susjednih segmenata, kako bi se životinja mogla prevrnuti i postići formu kugle.[3][4] Svaka metamera je ventralno imala međusobno slične noge, kao i na i glavi.
Pigidij uredi
Pigidij je činio završni dio tijela i sastojao se od promjenljivog broja stopljenih metamera koje su tvorile kompaktnu strukturu. Svaki segment je nosio par nogu sličnih onima na glavi i grudnom košu; ponekad je bilo nekoliko višečlanih vršnih cerki.
Dodaci uredi
Noge trilobita pokazuju iznenađujuće ujednačenost. Obično se smatra da su imale ulogu škrga i za veslanje, te koks, iz kojeg polazi endopodij, sastavljen od šest članaka i jednog terminalnog. Udovi su bili zaštićeni bočnim izbočenjima zvanim "pleuralni režnjevi", koji su se pružali prema središnjem aksijalnom režnju.
Ove vrste dodataka nisu homologni organi sa dodacima drugih skupina člankonožaca. Kod rakova, koji imaju i prstenasti dodatak, vanjska grana (egzopodij) je umetnuta u koku. Ni oni nisu direktno povezani sa dodacima kod Merostomada.
Spolni dimorfizam uredi

Izuzetan vremenki niz zabilježen u Murerovim kambrijskim naslagama i obilje fosila trilobita omogućili su, 2003. godine, identifikaciju apolnog dimorfizma kod trilobita, sa datiranjem najstarijieg slučaja spolnog dimorfizma poznatog u životinjskom carstvu. Za to su odabrane četiri filogenetsko bliske vrste sa širokom stratigrafskom rasprostranjenošću: Acadoparadoxides mureroensis, Eccaparadoxides rouvielli, E. sequeirosi i E. mediteranski. U svim tim slučajevima utvrđeno je da su kod svake vrste postojala dva različita morfotipa i da se ta razlika zadržavala tokom sedimentne sukcesije ili, što je ekvivalentno, tokom vremena.[7] Obilježja koja se razlikuju ova dva morfotipa su različita su za svaku vrstu, ali uključuju i dužinu pigidija, manju kod mužjaka i varijacije u dužini nekih segmenata toraksa ženki, koji predstavljaju uže područje prsnog koša, što odgovara segmentima nakon onog u kojem bi se vjerojatno nalazila genitalna pora (drugi toraksni segment, kao članaka u danšnjih Xiphosura).[8]
Paleobiologija uredi
Razvoj trilobita uključivao je niz stadija larve. Najprimitivnije vrste imale su kompletan razvoj larve dok je u kasnijim izvedbama proces bio pojednostavljen.
Prvi stadij larve poznat je pod nazivom protaspis, koji u osnovi formiraju cefalon i protopigidij. Nakon toga uslijedila je faza meraspis u kojoj su neki segmenti toraksa i pigidija već bili diferencirani. Treće razdoblje ili holaspis uključuje larve koje su već imale potpunu metamerizaciju, ali još uvijek mnogo manju nego kod odraslih, što je postizano nakon niza presvlačenja.
Paleoekologija uredi
Trilobiti su bili isključivo morski organizmi, potpuno odsutni u slatkim i bočatnim vodenim okruženjima; zbog spljoštenog oblika, očiju u dorzalnom položaju i tvrdoće dorzalne površine proizlazi da je većina njih bila bentoska. Sigurno su mikrofagi, koji su filtrirali blato s dna u kojem su živjeli kako bi dobili hranu, jer im nije nedostajalo pribora za iskopavanje ili sličnih dodataka, kao ni drobilica. Neke vrste sekundarno su postale pelagijske. Bodlje su bile prisutne i u larvama zvanim protaspis, pa se stoga smatraju pelagijskim oblicima s velikim kolonizacijskim potencijalom.
Smanjenje i gubitak očiju različitih vrsta zasigurno je povezan s prilagodbom na afotičku zonu i kolonizaciji dubokom moru.
Tafonomija uredi
Kao posljedica presvlačenja, vrlo su česti nalazi disartikuliranih fosilnih Exuvia , posebno cefalona ili njegovi zasebni dijelovi i obrazi, kao i pigidij (ista jedinka može imati višestruke pokazatelje tokom postojanja).
Tragovi fosilnih orgaizama - Iknofosili uredi


Biološkoj aktivnosti trilobita pripisuju se, uglavnom, tragovi fosila opisanih kao Cruziana, Diplichnites i Rusophycus, koji su, s druge strane, drugačiji organizmi.[9] Obično se nalaze kao protuteža, u obrnutom reljefu, u dnu slojeva pješčenjaka ili kvarcita
- Cruziana je trag koji se tumači se kao fosilna staza uslijed kretanja i/ili ishrane (Repichnia i Pascichnia ) trilobita na pješčanom dnu, a uključuje, s obzirom na veliku raznolikost trilobita, brojne vrste. Ti uzdužni tragovi formirani su od dva žlijeba koji napuštaju središnji greben, ponekad s dva druga manja bočna utora, a u hiperaktivnih poput dva uzdužna grebena s središnjim utorom. U oba se slučaja, ostavili su sitne pridnene pruge u obliku "V", koje ukazuju na suprotni smjer kretanja prema naprijed pomoću formiranih dodataka. Dobili i neformalni naziv „Bilobites“, koji je sada u upotrebi.[10]
- Diplihniti se interpretiraju kao tragovi brzog premještanja (Repichnia ) na pješčanoj površini. Čini se da su dvije paralelne staze formirane mnoštvom malih otisaka usko udaljenih jedan od drugog. Sitni tragovi odgovaraju dodacima organizama, a njihova dubina i rasprostranjenost ukazuju na uvjete ovog pomjeranja. Ponekad se tragovi pojavljuju u obliku finih pruga, kada je organizam imao bočno pomicanje zbog morskih struja, a ponekad se razvija prema stazama tipa cruziana kroz prijelazne forme[11]
- Rusofikus shvata se kao trag odmarajućeg stopala ("cubichnia") ili možda hranjenja. Prikazane su kao dvije kratke i fino urezane brazde, dublje od tioa cruziana i obično izolirane od drugih staza. Oni su rezultat procesa iskopavanja ili ukopavanja organizma za prikrivanje ili zarivanje u kojem su označeni tragovi dvije režnjevite pruge, formirane pomicanjem dviju skupina dodataka.[12]
Prvi trilobiti uredi
Najstariji poznati fosilni zapisi trilobita poznati su za Fallotaspidoidea i Redlichioidea, oba reda Redlichiida i bigotinide (red Ptychoparida), natporodica Ellipsocephaloidea ), datirana od prije 540 do 520 miliona godina. Prvi trilobiti uključuju „Profallotaspis jakutensis“ (Sibir), Fritzaspis spp. (zapadni dio Sjedinjenih Država), Hupetina antiqu (Maroko). Smatra se da trilobiti potiču iz sadašnjeg Sibira, s naknadnin rasprostranjenjem i evolucijskom radijacijom iz ove regije.[13][14]
Galerija uredi
-

-

-
 Trilobita
Trilobita -

-




Također pogledajte uredi
Reference uredi
- ^ Brusca, R. C. & Brusca, G. J., 2005. Invertebrados, 2ª edición. McGraw-Hill-Interamericana, Madrid (etc.), XXVI+1005 pp. ISBN 0-87893-097-3.
- ^ Riba, O. & Reguant, S., 1988. Registre fòssil. Història Natural dels Països Catalans, 8. Enciclopèdia Catalana, S. A., Barcelona, 478 pp. ISBN 84-7739-022-3.
- ^ Scholtz Gerhard (2008): Scarab beetles at the interface of wheel invention in nature and culture? Contributions to Zoology, 77:3. National Museum of Natural History Naturalis.
- ^ http://www.ctoz.nl/vol77/nr03/a01 Arhivirano 21. 10. 2016. na Wayback Machine | issn = 1875-9866 |access-date = January 19, 2009.
- ^ Dies Álvarez, M. E.; Rushtonc, A. W. A.; Gozalo, R.; Pillola, G. L.; Liñán, E. y Ahlber, P. (2010) «Paradoxides brachyrhachis Linnarsson, 1883 versus Paradoxides mediterraneus Pompeckj, 1901: a problematic determination». GFF, 132: 95-104
- ^ Gozalo, R.; Andrés, J. A.; Chirivella, J. B.; Dies Álvarez, M. E.; Esteve, J.; Gámez Vintaned, J. A.; Mayoral, E.; Zamora, S. y Liñán, E. (2010) «Murero y la explosión del Cámbrico: controversias acerca de este acontecimiento Arhivirano 13. 11. 2013. na Wayback Machine». Enseñanza de las Ciencias de la Tierra, 18(1): 47-59
- ^ Gozalo, R.; Liñán, E. y Dies, M. E. (2003) «Intraspecific dimorphism in an evolutionary series of paradoxidids from the Middle Cambrian of Murero, Spain». En: Lane, P. D.; Siveter, D. J. y Fortey, R. A. (eds.) Trilobites and their relatives (proceedings of Oxford conference 2001). Special Papers in Palaeontology, 70: 141-156
- ^ Liñán, E.; Gámez Vintaned, J. A.; Gozalo, R.; Dies Álvarez, M. E.; Chirivella Martorell, J. B. y Esteve, J. (2009) «Las especies de "paradoxides" del yacimiento cámbrico de Murero y el hallazgo de dimorfismo sexual en los trilobites Arhivirano 10. 6. 2015. na Wayback Machine». Naturaleza Aragonesa, 23: 4-11
- ^ Luis A. Buatois, M. Gabriela Mángano (2011). Organism-Substrate Interactions in Space and Time. Cambridge University Press.CS1 održavanje: upotreba parametra authors (link)
- ^ KU Icnology (ured.). "Cruziana". Pristupljeno 18. 8. 2015.
- ^ KU Icnology (ured.). "Diplichnites". Pristupljeno 18. 8. 2015.
- ^ KU Icnology (ured.). "Rusophycus". Pristupljeno 18. 8. 2015.
- ^ Lieberman, B. S. (2002) «Phylogenetic analysis of some basal early Cambrian trilobites, the biogeographic origins of the eutrilobita, and the timing of the Cambrian radiation Arhivirano 5. 3. 2016. na Wayback Machine». Journal of Paleontology, 76(4): 692–708
- ^ Fortey, Richard (2000) Trilobite!: Eyewitness to Evolution. Londres: Harper-Collins ISBN 0-00-257012-2
Bibliografija uredi
- Liñán, E. (1996): Los trilobites. En: Melic, A. (Ed.): PaleoEntomología [sic]. Boletín de la Sociedad Entomológica Aragonesa, 16: 45-56
- Rábano, I. (1999): Historia evolutiva de los trilobites. En: Melic, A.; Haro, J.J. de; Méndez, M. y Ribera, I. (Eds.): Evolución y Filogenia de Arthropoda. Boletín de la Sociedad Entomológica Aragonesa, 26: 225-233
Vanjski linkovi uredi
| Commons ima datoteke na temu: Trilobiti |
- http://mdbourrie.googlepages.com/trilobites Arhivirano 6. 11. 2012. na Wayback Machine Mark Bourrie's trilobite collection]
- http://www.trilobites.info/ A Guide to the Orders of Trilobites]
- https://web.archive.org/web/20161021001258/http://www.ctoz.nl/vol77/nr03/a01]
- https://books.google.es/books?id=kztUtwymPIMC&lpg=PP1&hl=es&pg=PP1#v=onepage&q&f=false]
- https://web.archive.org/web/20090514045820/http://www.mundo-geo.es/viajes-y-expediciones/expediciones/hallan-el-yacimiento-con-los-mayores-trilobites-del-mundo